2018-10-05 東京大学
1. 発表者:
添田 翔(沖縄科学技術大学院大学 ポストドクトラルスカラー
/東京大学大学院総合文化研究科広域科学専攻 特任研究員:研究当時)
大杉 美穂(東京大学大学院総合文化研究科広域科学専攻 教授)
2. 発表のポイント:
◆マウスの卵には、受精時に精子の核を受精卵の核へと変換するための十分な時間を保証する 分子メカニズムが備わっていることが明らかになり、またそのメカニズムの詳細が解明され ました。
◆受精を成立させるために哺乳類に備わっている、他の脊椎動物とは異なる特有のしくみの1 つが明らかになりました。
◆本研究成果により、哺乳類の受精についての理解が進みヒトや畜産動物の不妊の原因解明や 克服につながることが期待されます。
3. 発表概要:
受精卵の中では、卵と精子がもっていた染色体から 1 つずつ核(雌性前核と雄性前核(注 1)) が作られます。多くの脊椎動物では受精開始から 30 分ほどで前核が作られるのに対し、哺乳 類では数時間かかるという特性があります。しかし、前核形成まで数時間かかるのはどのよう なしくみによるのか、またどのような重要性があるのかは明らかになっていませんでした。
東京大学総合文化研究科の添田翔特任研究員、大杉美穂教授らは、これまで卵での役割が哺 乳類でのみ不明であった RSK(p90 ribosomal S6 kinase)というタンパク質リン酸化酵素(注 2)が、哺乳類の受精卵特有の時間制御を担っていることを見出しました。また、マウス受精 卵の前核形成までの時間を人為的に短くしたところ、受精卵が分裂する際に精子由来の染色体 が正常に受け継がれないことが明らかになりました。
本研究により、哺乳類は精子の核を受精卵の雄性前核へと変換するために長い時間を必要と し、卵にはその時間を保証するしくみが備わっていることが示されました。受精は普遍的な生 命現象ですが、哺乳類は特有のしくみをもつことがわかり、ヒトや畜産動物の不妊の原因解明 につながることが期待されます。
4. 発表内容:
<研究の背景>
受精により卵と精子という生殖に特化した細胞が融合し、全能性をもつ受精卵が作られます。 脊椎動物の卵細胞は細胞分裂抑制因子(CSF)(注 3)のはたらきにより減数第二分裂(注 4) の中期(注 5)で停止して精子を待ちます(図 1)。精子との融合がきっかけとなり減数第二 分裂の後期へと進むと、分配された染色体のうち一組は第二極体(注 6)として受精卵外に放 出され、もう一組が核膜に包まれて雌性前核になります。一方、精子の核内 DNA にはプロタ ミンという精子特有の塩基性タンパク質が結合し高度に凝集していますが、卵細胞内に入ると核膜が消えてプロタミンが外され、DNA はヒストンに巻きついたヌクレオソーム構造(注 7) をつくり、雌性前核と同時に核膜に包まれて雄性前核になります。この過程は精子クロマチン (注 8)の再構築とよばれます。カエルなど多くの脊椎動物では受精開始から 30 分ほどで前核 が作られるのに対し、哺乳類では数時間かかるという特性があります(図 1)。しかし、前核 形成まで数時間かかるのはどのようなしくみによるのか、どのような重要性があるのかは明ら かになっていませんでした。
また、多くの動物種において CSF を構成する分子である RSK(図 1)は、マウス卵内では CSF としての機能をもたず、哺乳類の卵における RSK の役割は解明されていませんでした。
<研究内容>
核の形成にはタンパク質の脱リン酸化反応が必要であることから、本研究ではまず、さまざ まな脱リン酸化酵素を検討し、PP1 という脱リン酸化酵素をマウス卵に過剰に存在させること で前核形成までの時間を大幅に短縮することに成功しました。前核形成を早期化した影響は雌 性前核にはあまり見られませんでしたが、雄性前核には、①ヒストン量の減少、② 核の小型化、 ③DNA 損傷の増加、④第一卵割時の染色体分配の失敗、等の異常が高頻度に見られました(図 2)。本研究により、マウス精子クロマチンの再構築には、クロマチンが核膜に包まれること なく卵細胞内に 80 分ほど存在することが重要であることが示されました。
次に、この時間を確保するためのしくみの解明を目指し、マウス卵での RSK の役割に着目 しました。RSK の量や活性を低減させると前核形成が 1 時間ほど早まったことから、RSK が 前核形成のタイミングを遅くしていることがわかりました。では、RSK はどういった標的分子 をリン酸化することで前核形成を遅らせているのでしょうか。
細胞分裂期の序盤に消失した核膜は、染色体が娘細胞に分配されたあとで再び染色体の周囲 に作られ核が形成されますが、染色体が分配される前に核ができてしまうのを防ぐしくみがあ ります。分裂期にすべての染色体が整列するまで高い活性をもつリン酸化酵素 Cdk1 が、 MASTL または Greatwall(Gwl)と呼ばれるリン酸化酵素を活性化し、MASTL/Gwl の標的 分子を介して核形成に必要な脱リン酸化酵素 PP2A を抑制する、というシグナル伝達経路(注 9)です(図 3)。染色体の分配開始と同時に Cdk1 が不活性化されると、続いて MASTL/Gwl が不活性化され、最終的に PP2A が活性化して分配された染色体を核膜が包みます(図 3)。 本研究においてカエルとマウスの卵減数第二分裂の染色体分配開始から前核形成までの様子を 調べたところ、カエルでは Cdk1 の不活性化から 20 分以内に MASTL/Gwl が不活性化されて いましたが、マウスでは MASTL/Gwl の活性が 2 時間近く維持されており、これがマウス受精 卵での前核形成に数時間かかる要因であることがわかりました。
そこで次に RSK と MASTL/Gwl の関係に注目しました。哺乳類の MASTL/Gwl には RSK の 標的アミノ酸配列(注 10)が存在しており、実際に RSK がマウスの MASTL/Gwl をリン酸化 し活性化することが示されました。一方、カエルを始め哺乳類以外の動物種の MASTL/Gwl には RSK の標的アミノ酸配列が存在していませんでした。
以上のことから、受精開始後すぐに Cdk1 が不活性化されても、哺乳類では RSK によって MASTL/Gwl の活性が維持されるため、前核形成が受精開始から数時間後になる、というしく みが明らかになりました。
本研究により、哺乳類の卵には、全能性をもつ受精卵を作る上で欠かせない精子クロマチン の再構築を完遂できるよう、余裕をもって前核形成を遅らせるしくみが備わっていることがわ かりました(図 3)。このような受精を支える哺乳類特有のしくみは他にも存在し、基礎研究によりその詳細が解明されることで、ヒトや哺乳類畜産動物の不妊の原因解明や対処法の開発 につながることが期待できます。
本研究は東京大学大学院総合文化研究科広域科学専攻大杉研究室と道上研究室との共同研究 として実施されました。本研究は日本学術振興会および文部科学省の科学研究費助成事業、成 茂動物科学振興基金等の助成により支援されました。
5.発表雑誌:
雑誌名:「Developmental Cell」
論文タイトル:RSK-MASTL pathway delays meiotic exit in mouse zygotes to ensure paternal chromosome stability
著者:Shou Soeda, Kaori Yamada-Nomoto, Tatsuo Michiue, and Miho Ohsugi, *
6. 問い合わせ先:
東京大学大学院総合文化研究科広域科学専攻 教授 大杉 美穂(おおすぎみほ
7.用語解説:
(注 1)前核
受精卵内につくられる、ゲノム 1 セット分の染色体を含む核。卵由来の染色体からできる前核 を雌性前核、精子由来の染色体からできる前核を雄性前核という。
(注 2)タンパク質リン酸化酵素
タンパク質を構成するアミノ酸のうち、主にセリンやスレオニン、またはチロシンの側鎖に、 アデノシン三リン酸(ATP)からリン酸基を転移する酵素。キナーゼ。リン酸化されたタンパ ク質は構造や活性が変化することが多い。
(注 3)細胞分裂抑制因子(CSF)
未受精卵内に存在する因子で、脊椎動物では細胞分裂期を中期に停止させ続ける活性をもつ。 実態はリン酸化酵素 MOS とその下流にある MEK、ERK、RSK などの一連のリン酸化酵素、 およびその標的分子であり、Cdk1 の不活性化を阻害することで中期停止を引き起こす。
(注 4)減数分裂
動物においては精子、卵子という配偶子をつくるための細胞分裂。1 回の DNA 複製の後、2 回の連続した染色体分配・細胞質分裂(減数第一分裂、減数第二分裂)を行うことで、細胞の もつゲノムのセット数を半減させる。
(注 5)中期
細胞分裂の 1 つの時期であり、すべての染色体が紡錘体の中央部に整列している時期。整列し た染色体は複製後であり、中期につづく後期に入ると染色体は1組ずつ分配される。
(注 6)極体
動物の卵細胞は減数分裂の 2 回の分裂の際、大きな細胞と小さな細胞を生じる極端な不等割分 裂を行うが、減数第一分裂、第二分裂により生じた小さな細胞をそれぞれ第一極体、第二極体 と呼ぶ。極体はやがて消失し、受精卵形成や胚発生には関わらない。
(注 7)ヌクレオソーム構造
DNA がヒストン 8 量体複合体に 2 回転弱巻きついた構造。
(注 8)クロマチン
DNA とヒストンなどの DNA 結合タンパク質の複合体。
(注 9)シグナル伝達経路
1 つの細胞内で起こる情報伝達機構。あるシグナルを受けた分子が活性化されると、次の特定 の分子群を結合やリン酸化などの修飾を介して活性化し、またその分子が次の分子を活性化す るといった形で起こる一連の反応。
(注 10)リン酸化酵素の標的配列
あるリン酸化酵素が、基質特異性をもってリン酸基を転移するために必要な、リン酸化するア ミノ酸の周囲の特定の性質を示すアミノ酸配列。例えばRSKの標的配列はR/K-X-R/K-X-X-S/T (R:アルギニン、K:リジン、X:任意のアミノ酸、S:セリン、T:スレオニン。K/R は K または R を意味する)であり、最後尾の S または T をリン酸化する。
8.添付資料:

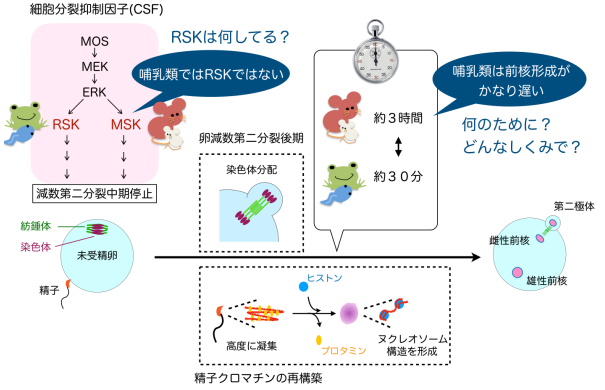
図 1 受精開始から前核形成までの過程の概要と研究開始時の疑問点
精子と卵が融合すると、CSF のはたらきにより減数第二分裂の中期で停止していた卵の細胞周 期は後期に進み、染色体分配と第二極体および雌性前核の形成が起こる。精子のクロマチンに は大規模な再構築が起こったあと雄性前核となる。哺乳類では前核形成までに数時間かかると いう特性があるが、どのようなしくみで、何のためにかは明らかになっていなかった。また、 多くの動物種において CSF を構成する分子である RSK は、マウス卵内では CSF としての機 能をもたず、哺乳類の卵での役割がわかっていなかった。
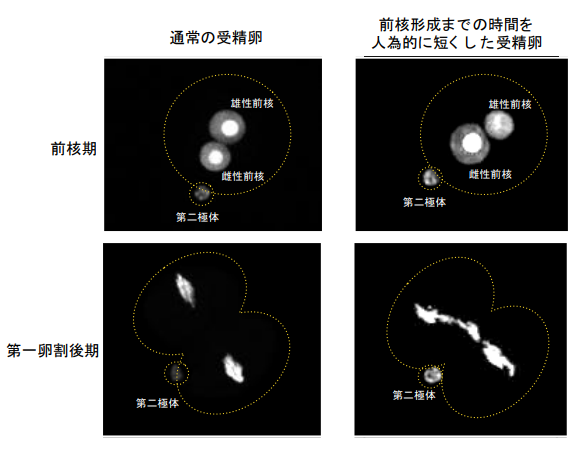
図 2 正常な受精卵と前核期形成タイミングを早期化した受精卵の比較
蛍光タンパク質を用い染色体を可視化した受精卵の蛍光顕微鏡画像。早期化受精卵では、前核 期の雄性前核が小さく、第一卵割後期において精子由来の染色体が正常に分配されない。黄色 点線は細胞の概形を示している。
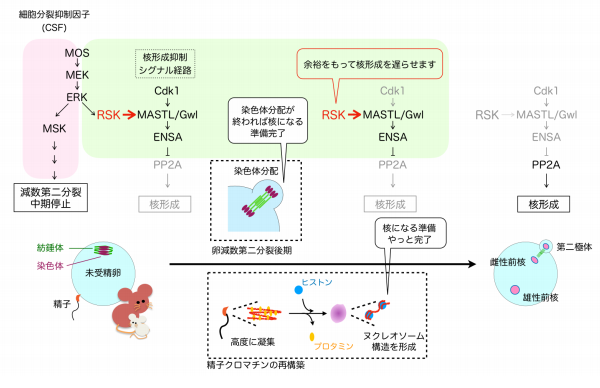
図 3 本研究成果の概念図
哺乳類以外の脊椎動物の卵減数第二分裂では、精子との融合後に Cdk1 の活性が失われると MASTL/Gwl もすぐに活性を失い核が形成されるが、哺乳類の卵では RSK が MASTL/Gwl を 数時間活性化し続けるため、核の形成が遅くなる。哺乳類の精子クロマチンの再構築には時間 がかかるため、核形成タイミングを遅くすることでその時間を保証している。