2021-09-02 東京大学
中山 北斗(生物科学専攻 助教)
発表のポイント
- 高速シークエンサー(注1)を用いて得られたゲノム情報を利用して、葉の形態が特徴的なトマトについて二つの原因遺伝子を同定し、そのうちの一つの遺伝子の進化の過程を明らかにしました。
- これらの結果から、特徴的な葉の形態を示すトマトについて、その成立過程と葉の発生メカニズムを明らかにしました。
- 本成果は、作物の形態進化に関して遺伝子レベルでの新たな事例を提示するとともに、作物などの形態の改良にも繋がることも期待されます。
発表概要
生物は多様な形態を示し、その中でも作物や家畜は、栽培化および家畜化、そしてその後の育種の過程で自然界では見られないような特殊な形態を示すことがあります。しかしながら、その過程で、どの遺伝子に、どのような変化が起きてそのような形態になったのかについては、依然として多くが不明のままです。
東京大学大学院理学系研究科の中山北斗助教、University of California, DavisのNeelima Sinha教授らの国際共同研究チームは、特定の地域のみで栽培され、特徴的な葉の形態を示すトマト(Solanum lycopersicum)の品種に着目し(図1)、その栽培の過程で、二つの異なるHOMEOBOX遺伝子(注2)に変化が起きたことで特徴的な葉の形態が生じたことを明らかにしました。
本研究により、作物の栽培の過程において、どの遺伝子に、どのような変化が起きたことで形態の多様化に繋がったのかが明らかになったとともに、今後、作物や観賞用の植物を用いた形態の改良に応用することが可能な知見が得られたと言えます。
この成果は、9月2日に「Current Biology」(online)に掲載されました。本研究は文部科学省科学研究費補助金、日本学術振興会科学研究費、NSF(米国科学財団)の支援を受けて行われました。
発表内容
生物は多様な形態を示します。その中でも作物や家畜は自然界では見られないような形態を示すことがあり、これは、栽培化やその後の育種の過程で、自然界では選択されないような形質が選ばれることがあるからだと考えられています。そのため、作物や家畜は、形態の多様性やその進化という観点から、非常に興味深い研究対象であると言えます。実際に、進化学に新しい展開をもたらしたチャールズ・ダーウィンは、150年以上も前から作物や家畜の多様性に着目しています。しかしながら、作物や家畜がどのような遺伝子にどのような変化を経て作出されてきたのかについては、その多くが不明なままです。
研究グループはこの問題にアプローチするために、heirloom tomato(注3)と呼ばれる、ある特定の地域でのみ栽培されてきたトマトに着目し、そのひとつである”Silvery Fir Tree”(SiFT)という品種を研究に用いました。このSiFTは一般的なトマトと比べて葉の複雑性が非常に高く(図1)、その特徴的な葉の形態のために、食用の実をつけるにも関わらず、庭園への植栽や景観植物としての利用もされるほどでした。

図1:トマトの葉の形態の比較
(左)一般的なトマトの葉の形態を示す系統であるM82。(右)高い複雑性を示すheirloom tomatoであるSilvery Fir Tree。同じ生育条件下で4週間育てた実生を用い、4枚目の葉を図に用いた。(Bar = 2 cm)。
そこでまず、特徴的な葉の形態を示すSiFTと一般的なトマトの葉の形態を示す系統であるM82を交配し、得られたF1世代(注4)を自家受粉させることでF2世代を得ました。このF2世代の葉は、SiFTのように葉の複雑性が高いものから、M82のようなものまでさまざまな形態が見られました。次に、高速シークエンサーを用いた遺伝学的解析を行い、F2世代の葉の複雑性が高い集団と低い集団について、それぞれの集団に特徴的な遺伝子領域を特定するBulked Segregant Analysisという解析を行いました。その結果、SiFTの2番染色体の一部の配列がM82と異なっており、その領域にあるBIPINNATA(BIP)というHOMEOBOX遺伝子に一塩基の欠失があることが明らかになりました。そこでWhole-mount in situ hybridizationという方法でBIP遺伝子の発現部位を調べると、発生中の葉原基の基部で発現していることがわかりました。この部位は小葉(注5)が発生する部位で、葉の複雑性と関係があります。次にBIP遺伝子の機能を破壊した変異体(bip変異体)の葉の形態を観察すると、M82と比べて複雑性が高くなっており、さらにその複雑性はSiFTと同等であることもわかりました。これらのことから、BIP遺伝子は葉原基の基部ではたらいて、葉の複雑性を抑制する機能を有しており、SiFTではBIP遺伝子の欠失変異のためにその機能が発揮できず、葉の複雑性が高くなっていると結論づけました。しかしながら、SiFTとbip変異体の葉を深層学習を用いた画像解析で比較してみると、その形態は同じではなく、SiFTの方が葉の小葉の幅が狭いこと、それ以外にも葉の内部構造の観察から、SiFTではbip変異体に比べて葉の維管束密度が低いことも明らかになりました。これらの結果は、SiFTの葉の形態に関与する遺伝子はBIP遺伝子だけではなく、別の遺伝子も関与していることを示唆しています。
次に、BIP遺伝子以外でSiFTの特徴的な葉の形態に関わった遺伝子を探索するために、SiFTとM82それぞれの茎頂や葉原基を用いたRNA-seq(注6)を行ない、そのデータを用いた共発現遺伝子ネットワーク(GCN)の比較解析を行ないました。その結果、モデル植物のシロイヌナズナ(Arabidopsis thaliana)においてWUSCHEL-RELATED HOMEOBOX 1(WOX1)として知られるHOMEOBOX遺伝子の相同遺伝子であるSlWOX1が、SiFTとM82のGCN間で最も異なっていることが明らかになりました。そこでこのSlWOX1の発現量を発生中の葉原基で調べたところ、SiFTではM82に比べて半分ほどになっていました。またSlWOX1遺伝子の発現部位を調べると、発生中の葉原基の周縁部で発現していることもわかりました。次に、SlWOX1遺伝子の機能を破壊した変異体の葉の形態を調べると、M82よりも小葉の幅が細く、葉内の維管束の密度が低いこともわかりました。そのため、本来は小葉の幅方向の成長や維管束密度の制御に促進的な機能をもつSlWOX1が、SiFTではその発現量が減ることではたらきが弱まり、M82に比べて細く、維管束密度の低い葉になっていることが示唆されました。また、bip変異体、slwox1変異体を交配した二重変異体では、葉の複雑性が増加し、小葉の幅は狭く、維管束密度は低いままでした。加えてbip変異体において、SlWOX1の発現量は変化しませんでした。したがって、これら二つの遺伝子の変化はお互いに干渉せず、その表現型は相加的であることも明らかになりました。以上のことから、SiFTでは、BIP遺伝子の変異による機能欠損とSlWOX1遺伝子の発現量の低下により、葉の複雑性が増加し、小葉の幅は狭く、そして維管束密度は低いという独自の葉の形態となったと結論づけました(図2)。
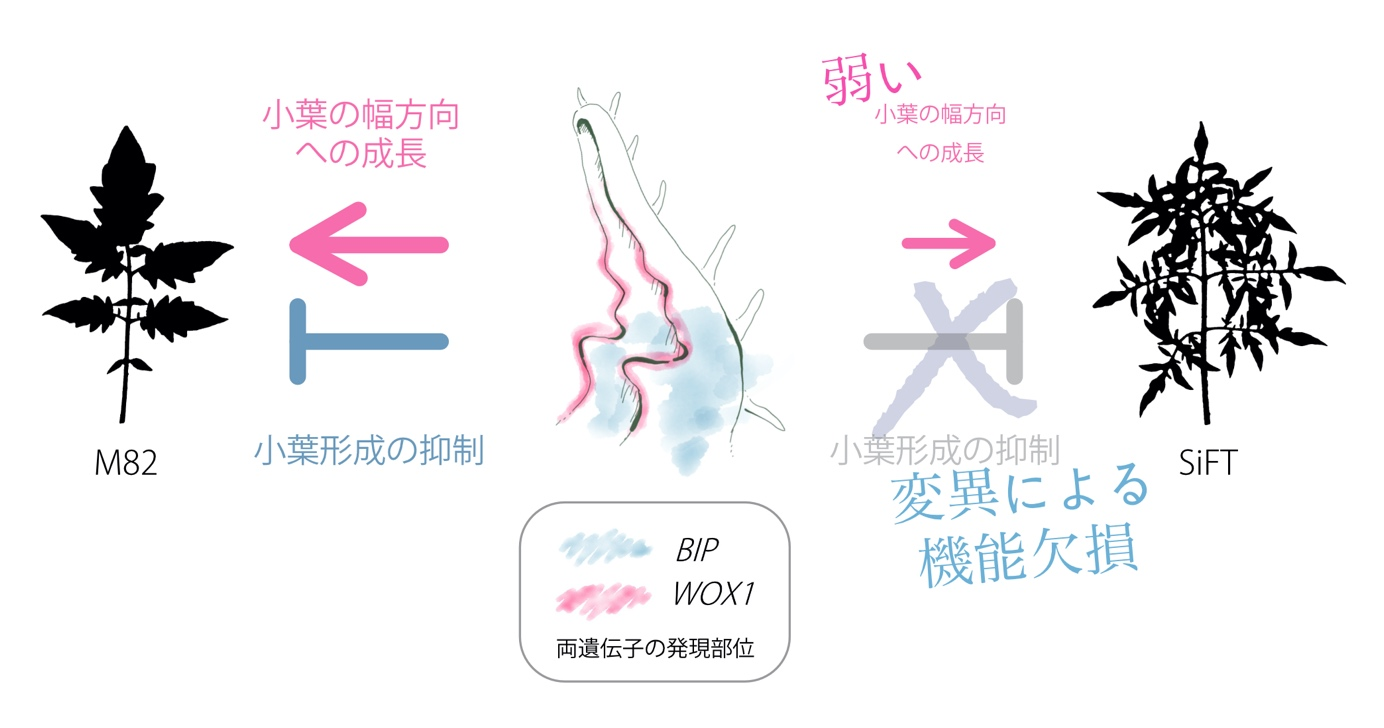
図2:Silvery Fir Treeの葉にみられる「複雑性が高く」そして「細い」葉になる仕組み
一般的なトマトにおいてみられる葉の形態はM82に代表されるような形をしており、これにはBIPやSlWOX1を含む数多くの遺伝子が関わっている。一方でSilvery Fir Treeでは、小葉の形成を抑制する機能をもつBIP遺伝子に変異が生じ、小葉形成を抑制できなくなっている。加えてそれぞれの小葉を幅方向に成長させるSlWOX1遺伝子のはたらきも弱くなっているために、「複雑性が高く」そして「細い」葉になる。
最後に研究グループは、SiFTに見られるbip変異が、進化の過程でどのように生じたのかについても調べました。これには、これまでにデータベースに登録されているさまざまなトマトのゲノムデータの他に、新たに複数のheirloom tomatoのゲノムシークエンスを行ない、それらのゲノムデータを用いて進化の過程を推定しました。その結果、解析に用いたトマトの中でSiFTに近縁なもの、SiFTの祖先系統との間で交雑が示唆されたもののなど、SiFTに関連のあるいずれの系統のゲノムからもbip変異は見つかりませんでした。実際にそれらのトマトの葉の形態はM82と同じく複雑性が低い形態であり、栽培化前のトマトの野生種も複雑性が低い形態であることが知られています。これらの事実から、SiFTに見られるbip変異はその栽培の過程で新規に獲得されたものであり、他のトマトから交雑などによって導入されたものではないことが示唆されました。
以上の解析から、heirloom tomato であるSiFTにみられる、複雑性が高く、小葉の幅が狭く、そして維管束密度が低いという特徴的な葉の表現型は、BIP遺伝子とSlWOX1遺伝子という二つのHOMEOBOX遺伝子の変化によってもたらされたことが明らかとなりました。また、その中でもbip遺伝子にみられる変異は、他のトマトからの導入によるものではなく、その栽培の過程で新規に獲得されたものであることが明らかになりました(図3)。
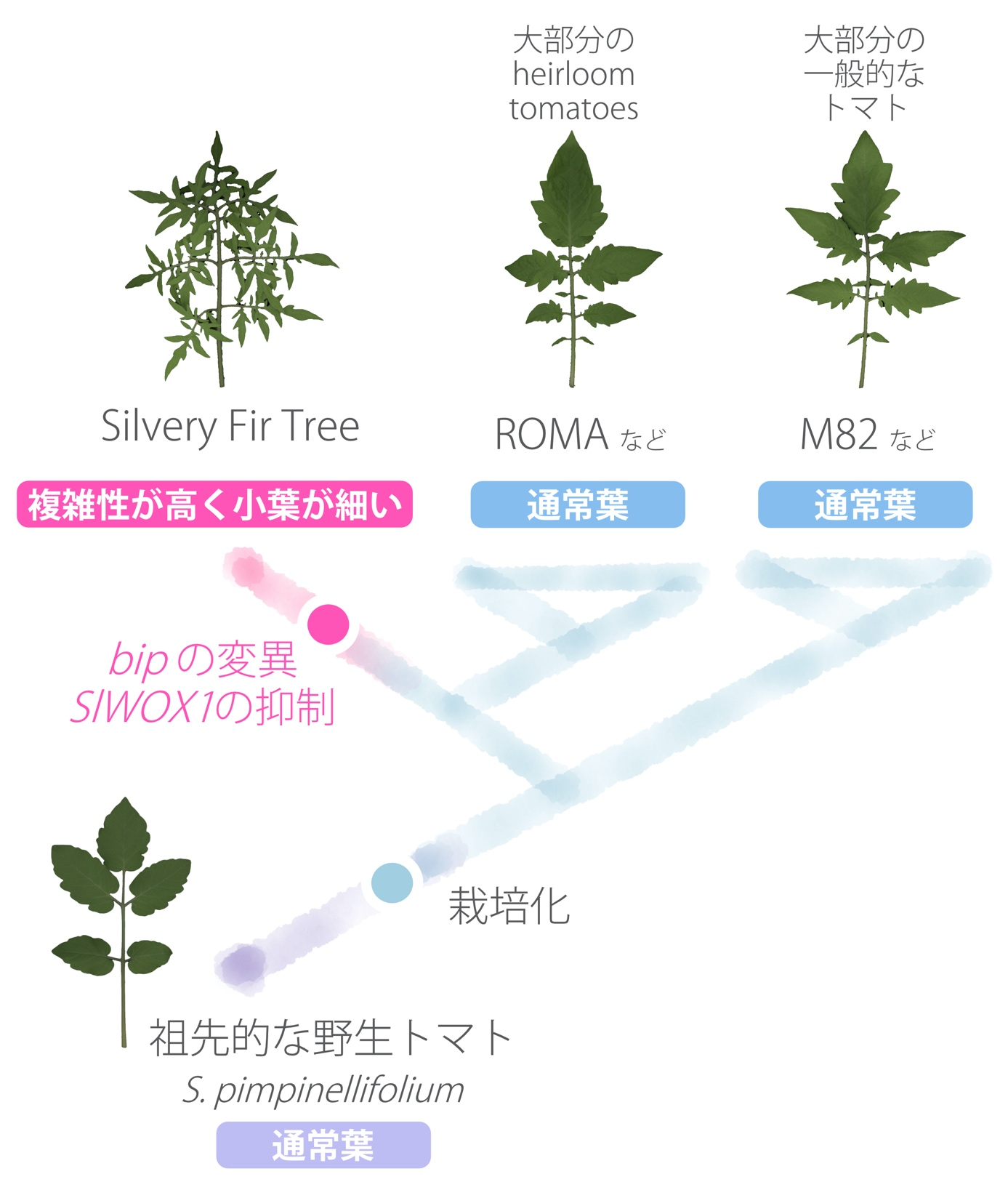
図3:トマトの栽培化や育種の歴史の中でみるSilvery Fir Treeの成立過程
大多数のトマトはM82と同じような葉の形態をしている。また、栽培化される前の野生トマトもM82でみられるような葉の形態をしている(Solanum pimpinellifoliumは現在のトマトの祖先種であることが知られている)。今回の研究から、Silvery Fir Treeの高い複雑性を示し、加えて小葉の幅が狭いという、他のトマトにはみられない特徴的な葉の表現型は、BIP遺伝子とSlWOX1遺伝子という二つのHOMEOBOX遺伝子の変化によってもたらされたことが明らかとなった。
葉は多くの植物において見られ、その形態はとても多様です。今回の場合のように、栽培の過程で特殊な形態が選抜され、私たちの目を楽しませてくれることもあります。加えて近年の研究では、葉の形態や維管束の密度が実の糖度との関連があることが報告されており、これらを改変することで実の食味や栄養素を改良することが可能になることも考えられます。そのため本研究の成果は、作物の栽培の過程において、どの遺伝子に、どのような変化が起きたことで形態の多様化に繋がったのかという進化的な観点からだけでなく、作物の品種改良にとっても重要な知見をもたらすものであると言えます。
発表雑誌
- 雑誌名
Current Biology論文タイトル
Leaf form diversification in an ornamental heirloom tomato results from alterations in two different HOMEOBOX genes著者
Hokuto Nakayama, Steven D. Rowland, Zizhang Cheng, Kristina Zumstein, Julie Kang, Yohei Kondo, Neelima R. SinhaDOI番号
10.1016/j.cub.2021.08.023論文URL
Redirectingdoi.org

用語解説
注1 高速シークエンサー
一度に数億~数十億塩基のDNAを解読することが可能な装置。これにより、従来に比べてゲノム解析や網羅的遺伝子発現解析を容易に行うことが可能になった。現在も精力的に改良が進んでおり、一度に解読可能な総塩基数や、リードと呼ばれる実際に得られる配列あたりの長さが増えている。
注2 HOMEOBOX遺伝子
ショウジョウバエのホメオティック突然変異の原因遺伝子に保存されていた配列(ホメオボックス)を持つ遺伝子。植物、動物、および菌類にみられ、発生過程の制御に関わることが知られている。
注3 heirloom tomato
ある特定の地域だけ、場合によっては家族内などでのみ何世代も栽培され、受け継がれて来たトマト。日本の京野菜などがイメージとしては近い。現在主に流通しているトマトとは異なり、味や色、形などが非常に多様であり、園芸や料理などにも好まれる。
注4 F1世代
雑種第一代のこと。異なる両親の交雑の結果生じた、第一世代目の子孫。雑種第二代の場合はF2となる。
注5 小葉
トマトのような、一枚の葉が複数の単位(小さい葉のようなもの)からなる葉を複葉と呼び、その一つひとつの単位を小葉と呼ぶ。小葉が多くなると葉の複雑性は高くなる。また、複葉に分類されない葉(葉縁部が明確に分かれていないもの)は単葉と呼ばれる。
注6 RNA-seq
高速シークエンサーを用いて、組織あるいは1細胞内の転写産物を解読し、網羅的に発現量の定量が可能な手法。さらにそのデータを統計的に処理することにより、似たような発現パターンをしている遺伝子を抽出し、それらの関係性をネットワークの形(共発現遺伝子ネットワークと呼ばれる)で表現することなども可能である。