2024-11-01 理化学研究所
理化学研究所(理研)生命医科学研究センター 免疫転写制御研究チームの谷内 一郎 チームリーダーらの国際共同研究グループは、転写因子[1]BCL11BをコードするBCL11B遺伝子のミスセンス変異[2]による先天性免疫異常症(IEI)[3]の病態を、モデルマウスの解析から明らかにしました。
本研究成果は、IEI病態のさらなる理解に貢献すると期待されます。
BCL11Bは免疫担当細胞であるT細胞の発生に重要な転写因子で、その変異はT細胞の分化や機能異常を伴うIEIの原因となります。
今回、国際共同研究グループは、IEIの患者に同定されたBCL11Bミスセンス変異がどのような病態を引き起こすのかをモデルマウスを用いて解明しました。
本研究は、科学雑誌『Nature Immunology』(11月1日付:日本時間11月1日)に掲載されました。

今回の研究の概要
背景
先天性免疫異常症(IEI)は、免疫機能に重要な役割を担う遺伝子の先天的な変異が原因となり発症します。免疫はウイルスや病原菌から体を守る大切な機能であり、その破綻は感染症や自己免疫疾患[4]へのリスクを高めます。IEIの原因となる変異はさまざまで、変異が起こった遺伝子や変異の種類によって病態は異なります。
骨髄では、さまざまな免疫細胞をつくる能力を持った造血前駆細胞がつくられ、この造血前駆細胞が胸腺へ移動すると、獲得免疫を担うT細胞に分化して他の免疫細胞への分化は抑制されます。このT細胞への分化を誘導する転写因子がBCL11Bです。BCL11Bは亜鉛フィンガー構造[5]を持ち、二量体[6]として亜鉛フィンガー構造を介してDNAに結合、遺伝子の発現を制御します。BCL11B遺伝子はT細胞の発生や機能に必要な遺伝子で、T細胞の減少症や欠損症を呈するIEI患者でその変異が報告されています。
本研究の共同研究者でもあるジェニファー・パック博士らのグループは2016年にT細胞欠損症を呈するIEI小児患者(患児)を報告しました注1)。報告の中で、その原因として同定されたのがBCL11Bの441番目のアミノ酸であるアスパラギン(N)がリジン(K)に置換されるN441Kミスセンス変異です。さらにパック博士らはN441K変異BCL11Bタンパク質が、正常なBCL11Bタンパク質の機能を阻害する優勢阻害効果[7]を有することを明らかにしています。
そこで、国際共同研究グループは、N441K変異BCL11Bタンパク質が引き起こす病態の解明に挑みました。
注1)D. Punwani, Y. Zhang, J. Yu, M.J. Cowan, S. Rana, A. Kwan, A.N. Adhikari, C.O. Lizama, B.A. Mendelsohn, S. P. Fahl, A. Chellappan, R. Srinivasan, S.E. Brenner, D.L. Wiest, J.M. Puck, Multisystem Anomalies in Severe Combined Immunodeficiency with Mutant BCL11B]
研究手法と成果
国際共同研究グループはヒトBCL11B遺伝子のN441Kミスセンス変異に相当するマウスBcl11b遺伝子のN440Kミスセンス変異を有する遺伝子改変マウス[8](N440K変異マウス)を作製しました。
表現型解析により、N440K変異マウスが小児患者(患児)と同様にT細胞減少症を示すことが分かりました。
そこでN440K変異新生児マウスの胸腺を解析したところ、T細胞への分化が遅延しており、さらに健常な胸腺には観察されない異常な細胞集団が出現することが判明しました。この異常な細胞は免疫細胞の一つであるナチュラルキラー(NK)細胞[9]と同じ特徴を持っていましたが、この細胞がNK細胞なのかNK細胞に似た別の細胞なのかは分かっていません。そのためここではこの異常な細胞をNK様細胞と表記します。
N440K変異Bcl11bタンパク質が、正常なBcl11bタンパク質の胸腺内でのT細胞分化誘導機能を阻害するのであれば、N440K変異マウスにはBcl11bタンパク質そのものは存在するが、Bcl11bがT細胞の発生を支持する機能は不在であると考えられます。この仮説が正しい場合、Bcl11bタンパク質が存在しない状況でもN440K変異マウスと同じ症状が現れると予測されます。しかしBcl11bを完全になくした遺伝子欠損マウス[10]の胸腺では、T細胞の分化は阻害されましたが、NK様細胞は出現しませんでした。この結果からN440K変異Bcl11bタンパク質は正常なBcl11bタンパク質の機能を阻害することでT細胞の分化異常を引き起こすと同時に、別のタンパク質に対しても阻害効果を発揮し、NK様細胞の出現を誘発するのではないかと仮説をたてました。
Bcl11b遺伝子が属するBcl11遺伝子ファミリー[11]には、Bcl11b遺伝子に加えてBcl11a遺伝子も属しています。胸腺内の造血前駆細胞ではBcl11aとBcl11bが共にタンパク質として発現しており、Bcl11bがBcl11aと直接的に相互作用することが実験的に確認できました。そこでN440K変異Bcl11bタンパク質はBcl11aの機能を阻害するのではないかと考え、Bcl11a遺伝子欠損マウスの解析を行ったところ、胸腺内にNK様細胞の出現が観察されました。この結果から、N440K変異Bcl11bタンパク質は正常なBcl11bタンパク質に加えてBcl11aタンパク質に対しても優勢阻害効果を発揮すること(ヘテロマー干渉阻害)、NK様細胞の分化抑制にはBcl11aが重要であることが示唆されました。
このことからBcl11bタンパク質はBcl11aタンパク質と協調することで胸腺内でのNK様細胞の発生を抑えていると考えられます。そこで国際共同研究グループではN440K変異マウスやBcl11a遺伝子欠損マウスでNK様細胞の発生を促す因子を突き止めるために、Bcl11bタンパク質と相互作用するタンパク質を網羅的に解析しました。その結果、転写因子Tcf1がBcl11bタンパク質と相互作用すること、そしてその相互作用はN440Kミスセンス変異によって減弱することを突き止めました。N440K変異マウスの造血前駆細胞は試験管内で培養するとNK様細胞へと分化しますが、人為的にTcf1遺伝子の発現を抑制するとNK様細胞の分化が抑えられることが分かりました。
BCL11B/Bcl11bはヒトでもマウスでも胸腺内のT細胞分化に必須のタンパク質です。今回の研究結果から、N440K変異Bcl11bタンパク質が正常なBcl11bの機能を阻害することでT細胞分化が阻害され、T細胞減少症を引き起こしたと考えられます。またN440K変異Bcl11bタンパク質がBcl11aの機能も障害することで、本来なら抑えられているはずの造血前駆細胞のT細胞以外の細胞への分化が誘導されることを明らかにしました。
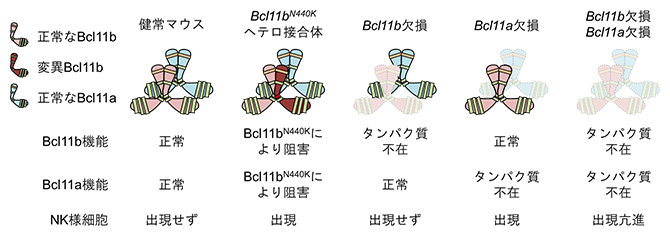
図 今回の研究で判明した結果のまとめ
N440K変異マウス胸腺内ではNK様細胞の異常な分化が観察されたが、Bcl11b遺伝子欠損マウス胸腺にNK様細胞は出現しない。一方でBcl11遺伝子ファミリーに属するBcl11a遺伝子の欠損マウス胸腺にNK様細胞が分化し、さらにBcl11bとBcl11aの両遺伝子を同時に欠損させることでその分化は亢進した。これらの結果から、胸腺内におけるNK様細胞の分化抑制にはBcl11bとBcl11aの両タンパク質の協調が重要であると考えられる。
今後の期待
変異タンパク質が同一遺伝子ファミリーに属するタンパク質に対して優勢阻害効果を発揮するヘテロマー干渉阻害は、谷内チームリーダーらが以前報告したIKZF3遺伝子のミスセンス変異でも観察された現象です注2)。今回のBCL11B遺伝子のN441Kミスセンス変異でも同様の現象が観察されたことは、ヘテロマー干渉阻害の病理学的な重要性を示唆するものです。さらに、本研究成果は、IEI病態の理解に貢献すると期待されます。
注1)2021年6月22日プレスリリース「新たな疾患発症メカニズムによる免疫不全症(AIOLOS異常症)の発見」
補足説明
1.転写因子
DNAに結合することで、遺伝子の発現を調節するタンパク質。
2.ミスセンス変異
遺伝子上のDNA配列が変化することで、翻訳されたタンパク質の一つのアミノ酸が別のアミノ酸に置換されてしまう変異。
3.先天性免疫異常症(IEI)
生まれながらの要因により免疫機能を担う要素(免疫担当細胞やタンパク質)の機能、あるいはそれらの要素そのものを欠如することで発症する免疫機能の異常による疾患。原発性免疫不全症(PID)ともいう。IEIはInborn Errors of Immunityの略。
4.自己免疫疾患
免疫機能の異常によって、本来は守られるべき自分自身の体に反応を示してしまう疾患。
5.亜鉛フィンガー構造
亜鉛イオンによって保持されるタンパク質上の特徴的な3次元的構造の一つ。転写因子に広く観察され、しばしばDNAとの結合に重要な役割を担う。
6.二量体
タンパク質2分子が結合し、一つの分子を形成したもの。同一のタンパク質で形成されるものをホモ二量体、2種類のタンパク質で構成されるものをヘテロ二量体と呼ぶ。
7.優勢阻害効果
変異タンパク質が正常なタンパク質の機能を阻害する効果。優勢阻害効果をもたらす変異を優勢阻害型変異と呼ぶ。
8.遺伝子改変マウス
学術研究の実施を目的に人工的に遺伝子を改変したマウス。
9.ナチュラルキラー(NK)細胞
がん細胞やウイルス感染細胞の排除に寄与する免疫細胞。
10.遺伝子欠損マウス
遺伝子改変マウスの一つで、任意の遺伝子が発現しないように遺伝子を改変したマウス。
11.遺伝子ファミリー
同一の祖先遺伝子から派生したために、塩基配列が高い相同性を共有する遺伝子群のこと。
国際共同研究グループ
理化学研究所 生命医科学研究センター
免疫転写制御研究チーム
チームリーダー 谷内 一郎(タニウチ・イチロウ)
研究員 奥山 一生(オクヤマ・カズキ)
研修生(研究当時)山下 基(ヤマシタ・モトイ)
(現 基礎科学特別研究員)
テクニカルスタッフⅡ(研究当時)太野(大石)路子(オオノ・(オオイシ)・ミチコ)
上級研究員(研究当時)海老原 敬(エビハラ・タカシ)
上級研究員(研究当時)香城 諭(コウジョウ・サトシ)
研修生(研究当時)サミュエル・J.H.・マーフィー(Samuel J.H. Murphy)
融合領域リーダー育成プログラム
上級研究員(研究当時)吉田 英行(ヨシダ・ヒデユキ)
(現 免疫転写制御研究チーム 客員研究員)
上級研究員(研究当時)イボ・ウー(Yibo Wu)
(現 粘膜免疫研究チーム 客員研究員)
粘膜システム研究チーム
チームリーダー 大野 博司(オオノ・ヒロシ)
専任研究員(研究当時)佐藤 尚子(サトウ・ナオコ)
(現 空間免疫制御理研ECL研究ユニット 理研ECLユニットリーダー)
東京医科歯科大学 発生発達病態学分野
教授 森尾 友宏(モリオ・トモヒロ)
ウルム大学(ドイツ)Institute of Molecular and Cellular Anatomy
大学院生 アルテミス・コウモンドウロウ(Artemis Koumoundourou)
助教授 クリストフ・ヴィグレフェッー(Christoph Wiegreffe)
教授 シテファン・ブリッシュ(Stefan Britsch)
カルフォルニア大学(米国)Department of Pediatrics
教授 ジェニファー・パック(Jennifer Puck)
ハッケンサック大学医療センター(米国)Center for Discovery and Innovation
研究員 シン・ジャオ(Xin Zhao)
教授 ハイ-ホゥェイ・シュエ(Hai-Hui Xue)
研究支援
本研究は、理化学研究所生命医科学研究センターPIDプロジェクト(研究代表者:谷内一郎)により実施し、日本学術振興会(JSPS)科学研究費助成事業基盤研究(C)「BCL11B変異によるヒト原発性免疫不全症発症機序の解明(研究代表者:奥山一生)」の助成を受けて行われました。
原論文情報
Kazuki Okuyama, Motoi Yamashita, Artemis Koumoundourou, Christoph Wiegreffe, Michiko Ohno-Oishi, Samuel J.H. Murphy, Xin Zhao, Hideyuki Yoshida, Takashi Ebihara, Naoko Satoh-Takayama, Satoshi Kojo, Hiroshi Ohno, Tomohiro Morio, Yibo Wu, Jennifer Puck, Hai-Hui Xue, Stefan Britsch, and Ichiro Taniuchi, “A mutant Bcl11bN440K protein interferes with Bcl11a function during T lymphocyte and neuronal development”, Nature Immunology, 10.1038/s41590-024-01997-5
発表者
理化学研究所
生命医科学研究センター 免疫転写制御研究チーム
チームリーダー 谷内 一郎(タニウチ・イチロウ)
報道担当
理化学研究所 広報室 報道担当
")
