
2023-01-12 理化学研究所,東京薬科大学,微生物化学研究会
理化学研究所(理研)環境資源科学研究センター ケミカルゲノミクス研究グループの伊藤 昭博 客員主管研究員(科技ハブ産連本部 創薬・医療技術基盤プログラム プロジェクトリーダー、東京薬科大学 生命科学部 教授)、吉田 稔 グループディレクター、東京薬科大学 生命科学部の高瀬 翔平 特別研究員(PD)、微生物化学研究会 微生物化学研究所の川田 学 部長らの共同研究グループは、ハイスループットスクリーニング[1]から得られたヒット化合物を基に合成した1,000種類以上の誘導体の構造活性相関[2]研究により、ヒストンメチル化[3]酵素G9aに対する新しい阻害剤「RK-701」の開発に成功し、RK-701によって発現するBGLT3長鎖ノンコーディングRNA[4](BGLT3 lncRNA)が鎌状赤血球症[5]患者の胎児型ヘモグロビン(HbF)[6]の誘導に普遍的な役割を持つことを発見しました。
本研究成果は、鎌状赤血球症の新しい治療薬の開発に貢献すると期待できます。
今回、共同研究グループは、特異的かつ低毒性で強力な新しいG9a阻害剤RK-701を開発しました。遺伝性疾患である鎌状赤血球症の治療戦略として、胎児期に存在し、出生後失われるHbFを再誘導させるという考え方が提唱されています。RK-701は、既存の鎌状赤血球症治療薬であるヒドロキシウレアよりも高効率にHbFの発現を誘導しました。さらに、BGLT3 lncRNAがRK-701だけでなく、ヒドキシウレアを含む他の誘導剤によるHbFの再活性化に重要な役割を果たすことを突き止めました。
本研究は、オンライン科学雑誌『Nature Communications』(1月12日付)に掲載されました。

G9a阻害剤RK-701による鎌状赤血球症治療戦略
背景
ヘモグロビンは血液中で酸素を運搬する「運送屋」として機能する、αグロビン鎖[7]2分子と非αグロビン鎖[7]2分子から構成される4量体タンパク質です。ヒトでは出生後に、非αグロビン鎖としてγグロビン鎖を持つ「胎児型ヘモグロビン(HbF、α2γ2)」から、βグロビン鎖を持つ「成人型ヘモグロビン(HbA、α2β2)」への切り替え(グロビンスイッチング)が起こります。
鎌状赤血球症は、βグロビン遺伝子(HBB)の点変異[8]によりHbAが異常なヘモグロビンS(HbS)に変化することで発症し、溶血性貧血や血管閉塞性クリーゼを主症状とする遺伝性疾患です。赤血球の形が鎌状になることから、この疾患名が付けられました。
鎌状赤血球症の治療戦略として、出生後に発現が抑制されるHbFを再誘導するという方法が提唱されています。実際、HbF誘導活性を示すヒドロキシウレアが鎌状赤血球症治療薬として臨床の場で使用されていますが、DNAの合成を阻害するため安全性と有効性に問題があります。全世界で毎年約30万人の新生児が鎌状赤血球症を発症し、非常に多くの患者が存在することから、より強力で安全なHbF誘導薬の開発が強く求められています。
そのような背景の下、ヒストンメチル化酵素G9aの阻害剤UNC0638がHbF誘導活性を持つことが報告され、鎌状赤血球症の治療標的としてG9aが有望であることが分かりました注1)。G9aは、ヒストンH3の9番目のリジン残基のジメチル化(H3K9me2)を触媒する酵素です。しかし、UNC0638を含めた既存のG9a阻害剤は毒性が強く、治療薬として使用することはできません。また、G9aによる胎児型γグロビン遺伝子の発現抑制のメカニズムはよく分かっていませんでした。
注1)EHMT1 and EHMT2 inhibition induces fetal hemoglobin expression. Blood. 2015 Oct 15;126(16):1930-9. doi: 10.1182/blood-2015-06-649087
研究手法と成果
共同研究グループは、まず約14万の化合物からG9a阻害活性を持つ化合物の探索を目的としたハイスループットスクリーニングを行いました。得られたヒット化合物とG9aの複合体の構造情報を基に化合物の最適化を行い、G9a阻害活性がヒット化合物よりも約200倍も高い「RK-701」の開発に成功しました(図1)。
RK-701は、関連するG9a以外のメチル基転移酵素に対して1,000倍以上の選択性を持ち、既存のG9a阻害剤と比較して著しく毒性が低く、薬物動態も良好でした。そこで、ヒト赤血球細胞に投与したところ、RK-701は胎児型γグロビンの発現を誘導し、既存の鎌状赤血球症治療薬ヒドロキシウレアよりも低濃度で優れた薬効を発揮することが分かりました。また、マウスにRK-701を投与すると、ヒトの胎児型γグロビンに相当する胚型εyグロビンの発現が誘導され、RK-701は動物モデルでも有効であることが分かりました。
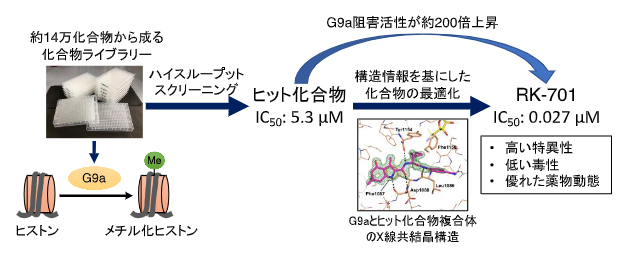
図1 G9a阻害剤RK-701の開発
約14万の化合物からG9aのヒストンメチル化酵素活性を阻害する化合物を探索。得られたヒット化合物とG9aの複合体のX線共結晶構造を基に化合物の最適化を行い、G9a阻害活性がヒット化合物より約200倍上昇したRK-701の創製に成功した。
次に、RK-701を用いて、G9aによる胎児型γグロビン遺伝子の発現抑制機構を調べました。RK-701により発現が増加する遺伝子を網羅的に調べたところ、数ある胎児型γグロビン発現制御因子の中で、BGLT3遺伝子の発現だけが非常に増加していることが分かりました。BGLT3は、胎児型γグロビン発現誘導活性を持つ長鎖ノンコーディングRNA(IncRNA)です。細胞内のBGLT3(BGLT3 lncRNA)の発現を低下させると、RK-701による胎児型γグロビンの発現誘導がほぼ完全に抑制されました(図2右上)。これらの結果から、G9aによる胎児型γグロビン遺伝子の発現抑制にBGLT3が重要な役割を果たしていることが分かりました(図2)。
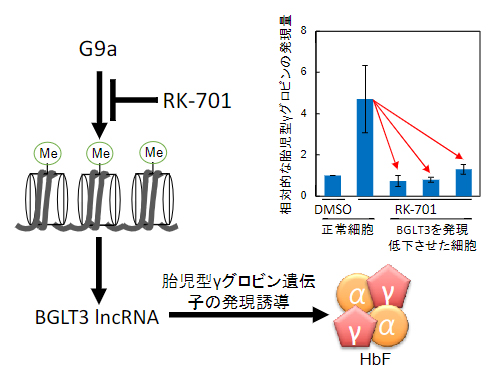
図2 BGLT3 IncRNA依存的なRK-701によるHbF誘導
ヒト赤芽球細胞株HUDEP-2細胞において、溶媒として用いたDMSO処理と比較して、RK-701処理により胎児型γグロビンの発現が増加した。RNA干渉法によりBGLT3(BGLT3 lncRNA)の発現を低下させると、このRK-701による胎児型γグロビンの発現増加はほぼ完全に抑制された(右上のグラフ)。G9aによる胎児型γグロビン遺伝子の発現抑制に、BGLT3が重要な役割を果たしている。
さらに、G9aによるBGLT3発現抑制機構を調べました。BCL11AとZBTB7Aは、胎児型γグロビン遺伝子の主要な転写抑制因子[9]として働くことが知られています。これらの転写抑制因子を欠損させると、(1)BGLT3の発現が高まりRK-701によるBGLT3の発現増加が見られなくなること、(2)BGLT3遺伝子座のH3K9me2レベルが低下することから、これらの転写抑制因子とG9aは協調してBGLT3の発現を抑制しているらしいことが分かりました。さらに(3)二つの転写抑制因子はG9aと相互作用してG9aをBGLT3遺伝子座付近に動員することによりH3K9me2の上昇を誘導し、BGLT3の発現を抑制することが示唆されました。
一方、G9aによるBGLT3遺伝子座付近のH3K9me2の増加は、H3K9me2結合タンパク質であるCHD4を介してBGLT3遺伝子座付近へのBCL11AおよびZBTB7Aの動員を増加させました。以上の結果から、これらの転写抑制因子がG9aをBGLT3遺伝子座付近へ誘導して近傍のH3K9me2を増加させ、それがCHD4を介してBGLT3遺伝子座付近に転写抑制因子を動員させH3K9me2がさらに増加するというH3K9me2が増幅される仕組みがBGLT3の発現抑制、ひいては胎児型γグロビン遺伝子の発現抑制の分子基盤であることが示されました(図3)。
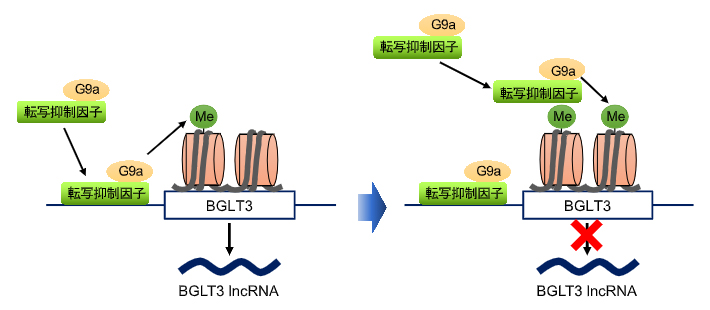
図3 G9aによるBGLT3(BGLT3 lncRNA)発現抑制機構
BCL11AとZBTB7Aという2種類の転写抑制因子がBGLT3遺伝子近傍に結合すると、それらが相互作用するG9aをBGLT3遺伝子近傍に呼び寄せる。すると、G9aによりBGLT3遺伝子座のヒストンH3がメチル化される(左)。メチル化されたヒストンH3は、転写抑制因子複合体に含まれるCHD4と相互作用することで、さらに転写抑制因子をヒストン領域に動員し、その結果G9aによるヒストンH3メチル化が増大する(右)。こうしてBGLT3遺伝子座は次々とメチル化が拡大され、その遺伝子発現が抑制されると考えられる。
既存薬であるヒドロキシウレアに加えて、DNAメチル化酵素[10]の阻害剤やヒストン脱アセチル化酵素[11]の阻害剤も、胎児型γグロビンの発現を増加させることが知られています。胎児型γグロビン発現誘導におけるBGLT3 lncRNAの重要性は、RK-701によるものだけにとどまりませんでした。つまり、ヒドロキシウレアを含めたこれらの阻害剤もBGLT3の発現を増加させること、BGLT3の発現を低下させると胎児型γグロビンの発現増加が抑制されることを明らかにしました。胎児期に発現するγグロビン遺伝子は、霊長類の中でもヒトを含む真猿類[12]に存在していますが、より原始的な原猿類[13]には存在しません。同様に、BGLT3遺伝子も真猿類に存在しますが、原猿類には存在しないことから、胎児型γグロビン遺伝子とBGLT3遺伝子は協調的に進化してきた可能性があります。以上の結果から、BGLT3は高等動物が進化上獲得したHbFの普遍的な活性化因子であると考えられます。
今後の期待
本研究では、新しい化学構造を持ち、特異的かつ強力なG9a阻害剤RK-701の開発に成功しました。RK-701は、UNC0638などの一般的なG9a阻害剤よりも毒性が著しく低く、既存の治療薬であるヒドロキシウレアよりも優れた安全性と有効性を示したことから、鎌状赤血球症治療薬として大いに期待できます。
また、本研究により、BGLT3 lncRNAが胎児型γグロビン遺伝子発現の普遍的な活性化因子であることが明らかになりました。RK-701だけでなく、安全にBGLT3 IncRNAの発現を高める方法を見いだすことができれば、鎌状赤血球症の新たな治療薬の開発につながると期待できます。
本研究成果は、国際連合が2016年に定めた17項目の「持続可能な開発目標(SDGs)[14]」のうち、「3.すべての人に健康と福祉を」に貢献するものです。
補足説明
1.ハイスループットスクリーニング
スクリーニングとは、目的の活性を持つ化合物を選別すること。ハイスループットスクリーニングとは、ロボットなどを用いて膨大な数の化合物から有用なものを迅速かつ高効率で選別するスクリーニングのこと。
2.構造活性相関
化合物の構造と生物学的な活性の関係のこと。構造活性相関を調べることにより、生物活性の高い化合物の設計が可能になる。
3.ヒストンメチル化
真核細胞では、DNAはヒストンと呼ばれるタンパク質に巻き付いて核内に収納されている。ヒストンタンパク質のリジンまたはアルギニン残基にメチル基が付加することをメチル化と呼ぶ。ヒストンがメチル化されることで、DNAの巻き付き方が変化し、それにより遺伝子の発現が変化する。H3K9me2は、ヒストンH3の9番目のリジン残基にメチル基が二つ付加したもので、遺伝子発現に抑制的に働く。
4.長鎖ノンコーディングRNA
ノンコーディングRNAは、タンパク質へ翻訳されずにRNAとして機能する分子の総称。長鎖ノンコーディングRNAは200ヌクレオチド以上から成るノンコーディングRNAを指す。
5.鎌状赤血球症
ヘモグロビンを構成するβグロビン遺伝子に変異が生じる遺伝性疾患の一つ。正常なヘモグロビンと比べ、体内に酸素を運ぶ機能が低下するために貧血症状などを呈する。
6.胎児型ヘモグロビン(HbF)
胎児期に存在するヘモグロビンのこと。胎児型ヘモグロビンはαグロビン2分子とγグロビン2分子から構成される。出生後、γグロビン遺伝子の発現が抑制され、代わりにβグロビン遺伝子が発現し、αグロビン2分子とβグロビン2分子から構成される成人型ヘモグロビンに置き換わる。
7.αグロビン鎖、非αグロビン鎖
どちらもヘモグロビンの構成因子。ヘモグロビンはαグロビン2分子と成人型のβグロビンまたは胎児型のγグロビン2分子から構成されるため、βまたはγグロビンを総称して非αグロビン鎖と呼ぶ。
8.点変異
正常なDNA配列が1塩基レベルで異なる配列に置き換わること。変異には置換以外に、塩基がなくなる欠失、余分に加わる挿入がある。本来のDNA配列から変化することで、翻訳されるタンパク質のアミノ酸配列も置換され、異常な活性を示す。
9.転写抑制因子
転写因子とは、特定のDNA配列に結合し、転写を調節するタンパク質。転写抑制因子は、転写抑制化に働く転写因子のこと。
10.DNAメチル化酵素
DNA配列中のシトシンにメチル基を付加する酵素。DNAメチル化は、遺伝子の発現を抑制する。
11.ヒストン脱アセチル化酵素
ヒストンなどのタンパク質のリジン残基に付加されたアセチル基を除去する酵素。ヒストンの脱アセチル化は遺伝子の発現を負に制御する。
12.真猿類
霊長類は大きく、真猿類と原猿類に分けられる。真猿類は、ヒトを含む高度に進化した哺乳類。真猿類はヒト、マントヒヒ、ニホンザルなどの狭鼻類とクモザル、オマキザルなどの広鼻類に二分される。
13.原猿類
真猿類と比較して原始的なサル類の総称で、キツネザル、メガネザルなどが分類される。一般的に樹上で生活し、小形で大脳の発達程度は低い。
14.持続可能な開発目標(SDGs)
持続可能な開発目標(SDGs)とは、2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標である。持続可能な世界を実現するための17のゴールから構成され、地球上の誰ひとりとして取り残さないことを誓っている。SDGsは発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に取り組んでいる。(外務省のホームページから一部改変して転載)
共同研究グループ
理化学研究所
環境資源科学研究センター
ケミカルゲノミクス研究グループ
客員主管研究員 伊藤 昭博(イトウ・アキヒロ)
(科技ハブ産連本部 創薬・医療技術基盤プログラム プロジェクトリーダー、東京薬科大学 生命科学部 教授)
グループディレクター 吉田 稔(ヨシダ・ミノル)
(創薬シード化合物探索基盤ユニット 基盤ユニットリーダー)
創薬シード化合物探索基盤ユニット
テクニカルスタッフⅠ 新真 由美(アラタ・マユミ)
テクニカルスタッフⅡ 中田 明子(ナカタ・アキコ)
技師 松岡 聖二(マツオカ・セイジ)
テクニカルスタッフⅠ 園田 健(ソノダ・タケシ)
創薬化学基盤ユニット
上級研究員 白井 文幸(シライ・フミユキ)
バイオリソース研究センター
細胞材料開発室
専任研究員 寛山 隆(ヒロヤマ・タカシ)
室長 中村 幸夫(ナカムラ・ユキオ)
生命機能科学研究センター
エピジェネティクス制御研究チーム
上級技師 丹羽 英明(ニワ・ヒデアキ)
技師 佐藤 心(サトウ・シン)
チームリーダー 梅原 崇史(ウメハラ・タカシ)
タンパク質機能・構造研究チーム
チームリーダー 白水 美香子(シロウズ・ミカコ)
脳神経科学研究センター 生体物質分析支援ユニット
専門技術員 臼井 正哉(ウスイ・マサヤ)
科技ハブ産連本部 創薬・医療技術基盤プログラム
研究嘱託 山口 時男(ヤマグチ・トキオ)
開拓研究本部 眞貝細胞記憶研究室
主任研究員 眞貝 洋一(シンカイ・ヨウイチ)
東京薬科大学 生命科学部
細胞情報科学研究室
特別研究員(PD)高瀬 翔平(タカセ・ショウヘイ)
助教 前本 佑樹(マエモト・ユウキ)
腫瘍医科学研究室
准教授 林 嘉宏(ハヤシ・ヨシヒロ)
教授 原田 浩徳(ハラダ・ヒロノリ)
微生物化学研究会 微生物化学研究所
第1生物活性研究部
部長 川田 学(カワダ・マナブ)
沼津支所
主任研究員 大石 智一(オオイシ・トモカズ)
研究員 大庭 俊一(オオバ・シュンイチ)
杏林製薬株式会社 わたらせ創薬センター 合成研究所
研究員 西ヶ谷 洋輔(ニシガヤ・ヨウスケ)
研究員 浪江 亮介(ナミエ・リョウスケ)
副主任研究員 角谷 龍展(スミヤ・タツノブ)
主任研究員 橋本 憲明(ハシモト・ノリアキ)
研究支援
本研究は、理研創薬・医療技術基盤プログラム「ヒストンメチル化酵G9aを標的としたβグロビン異常症治療薬の開発(プロジェクトリーダー:伊藤昭博)」、理研横断エピゲノム操作プロジェクト(研究代表者:眞貝洋一、伊藤昭博)、および脳神経科学研究センター生体物質分析支援ユニットによる支援を受けて実施し、日本医療研究開発機構(AMED)次世代がん研究シーズ戦略的育成プログラム「発がんに関わるヒストン修飾酵素を標的とした抗がん剤の開発(研究代表者:伊藤昭博)」、日本学術振興会(JSPS)科学研究費助成事業基盤研究(S)「革新的化学遺伝学による内在性代謝物の新機能の解明と応用(研究代表者:吉田稔)」、同若手研究「ヒストンメチル化酵素G9aによる胎児型ヘモグロビンの制御機構解析(研究代表者:高瀬翔平)」、同特別研究員奨励費「ヒストンメチル化酵素G9aを分子標的とした疾患治療法の開発(研究代表者:高瀬翔平)」による助成を受けて行われました。
原論文情報
Shohei Takase, Takashi Hiroyama, Fumiyuki Shirai, Yuki Maemoto, Akiko Nakata, Mayumi Arata, Seiji Matsuoka, Takeshi Sonoda, Hideaki Niwa, Shin Sato, Takashi Umehara, Mikako Shirouzu, Yosuke Nishigaya, Tatsunobu Sumiya, Noriaki Hashimoto, Ryosuke Namie, Masaya Usui, Tomokazu Ohishi, Shun-ichi Ohba, Manabu Kawada, Yoshihiro Hayashi, Hironori Harada, Tokio Yamaguchi, Yoichi Shinkai, Yukio Nakamura, Minoru Yoshida, Akihiro Ito, “A specific G9a inhibitor unveils BGLT3 lncRNA as a universal mediator of chemically induced fetal globin gene expression”, Nature Communications, 10.1038/s41467-022-35404-0
発表者
理化学研究所
環境資源科学研究センター ケミカルゲノミクス研究グループ
客員主管研究員 伊藤 昭博(イトウ・アキヒロ)
(科技ハブ産連本部 創薬・医療技術基盤プログラム プロジェクトリーダー、東京薬科大学 生命科学部 教授)
グループディレクター 吉田 稔(ヨシダ・ミノル)
東京薬科大学 生命科学部 細胞情報科学研究室
特別研究員(PD)高瀬 翔平(タカセ・ショウヘイ)
微生物化学研究会 微生物化学研究所 第1生物活性研究部
部長 川田 学 (カワダ・マナブ)
報道担当
理化学研究所 広報室 報道担当
東京薬科大学 総務部 広報課
微生物化学研究会 微生物化学研究所 知的財産情報部

