2022-05-19 理化学研究所
理化学研究所(理研)バイオリソース研究センター遺伝工学基盤技術室の越後貫成美専任技師、小倉淳郎室長(開拓研究本部小倉発生遺伝工学研究室主任研究員)、生命機能科学研究センター染色体分配研究チームの京極博久客員研究員、北島智也チームリーダーらの共同研究グループは、効率が著しく低かった一次精母細胞[1]を用いた顕微授精[2]技術の改良を行い、卵子の細胞質サイズを小さくすることによって、マウス産子の出生率の大幅な改善に成功しました。
一次精母細胞は減数分裂[3]前の雄性生殖細胞であり、多くの無精子症[4]男性にもその存在が認められていることから、本研究成果は将来的に新たな不妊治療法の一つとなる可能性があります。
一次精母細胞を用いた顕微授精法による産子作出は既に20年以上前にマウスで報告されていますが、その出生率はわずか数%です。この原因として、一次精母細胞注入後の卵子内での減数分裂において染色体異常が高頻度に生じることが挙げられていましたが、その改善方法は見つかっていませんでした。
今回、共同研究グループは、卵子の細胞質サイズを小さくすることで減数分裂中の染色体の動態が安定する現象に着目し、一次精母細胞の顕微授精に小さくした卵子を用いることを試みました。すると、通常サイズの卵子に注入した場合と比べて、小さい卵子中では減数分裂時の染色体の異常が大幅に軽減されました。実際に得られた顕微注入胚を母体へ移植したところ、出生率が約20倍まで向上しました。さらに一次精母細胞で精子発生が停止している無精子症マウスからの産子獲得にも成功しました。
本研究は、科学雑誌『EMBO Reports』オンライン版(5月19日付:日本時間5月19日)に掲載されました。

ライブイメージング解析による一次精母細胞注入卵子における減数分裂の観察
背景
ヒトや動物の生殖補助技術[5]として顕微授精技術が広く用いられており、近年、精子以外に減数分裂後の未成熟精子(精子細胞)からも産子が得られるようになってきました。さらにマウスでは、減数分裂前の一次精母細胞からも産子の作出に成功しています。しかし、一次精母細胞を用いて産子を得るには、一次精母細胞の染色体が複雑な減数分裂を卵子の中で完了しなければなりません(図1)。
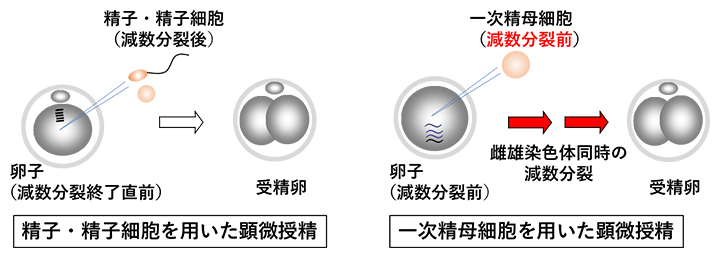
図1 精細胞の違いによる顕微授精後の細胞分裂の違い
減数分裂を終了している精子・精子細胞を顕微授精に用いる場合、排卵された成熟卵子(減数分裂終了直前)に注入することで受精が成立する。一方、一次精母細胞は減数分裂前の細胞なので、卵子の内部で減数分裂を進行させる必要がある。
このため、一次精母細胞を用いた顕微授精では染色体異常[6]を生じることが多く、その結果、出生率は非常に低いのが現状でした。一方、京極博久客員研究員と北島智也チームリーダーは、卵子はその巨大な細胞質サイズのために染色体分配異常を起こしやすいことを報告しています注)。
そこで本研究では、顕微操作により細胞質サイズを半分程度に減らしたマウス卵子を一次精母細胞の顕微授精に用いることで、注入胚の染色体異常が減少するか、そして胚移植後の出生率の向上につながるかを調べました。
注)2017年5月9日プレスリリース「卵母細胞はその大きさゆえに間違えやすい」
研究手法と成果
共同研究グループはまず、卵子の細胞質サイズと一次精母細胞の顕微授精後の染色体異常の関係を解析しました。その結果、細胞質サイズがそのままの卵子(コントロール卵子)では2%(1/59)のみが正常染色体を保持していたのに対し、細胞質サイズを1/3~1/2程度に減らした卵子では21%(13/62)が正常染色体を保っていました。観察された染色体異常の多くは、姉妹染色分体の早期分離[7]でした。さらに、一次精母細胞注入直後から減数分裂終了までの間の染色体の動態について、雌雄それぞれの染色体を識別してライブイメージング解析を行いました。その結果、大部分の染色体異常は精母細胞由来の染色体に生じており、それが卵子の細胞質サイズを小さくすることで改善されることが明らかになりました(図2)。
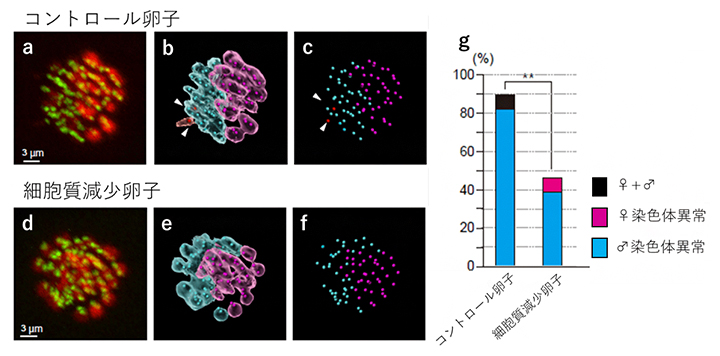
図2 ライブイメージング解析による一次精母細胞注入卵子における減数分裂の観察
抗体を使って染色体を赤色、染色体の中心にある動原体(紡錘体微小管の接着部位)を緑色に蛍光染色し、染色体の蛍光強度の違いから雄性(弱)と雌性(強)の染色体を識別した(a、d)。雄性染色体を青色、雌性染色体をピンク色に区別し(b、e)、それぞれの動原体位置を点描した(c、f)。コントロール卵子(上)では20ペアの雌性染色体、19ペアの雄性染色体、およびペアになっていない雄性染色体(赤点)が観察された。g:減数分裂期に染色体異常が生じている一次精母細胞注入卵子を数えたところ、コントロール卵子の9割近くが染色体異常を示したのに対し、細胞質減少卵子では異常が半分以下であった。また、異常染色体の大部分は雄由来であった(**P<0.01)。
次に、産子までの発生能の解析を行いました。2細胞期まで発生した一次精母細胞由来胚を偽妊娠雌マウスの卵管に移植したところ、コントロール胚では1%(1/96)の出生率(胚移植当たり)だったのに対し、細胞質減少卵子の胚では19%(17/90)と、20倍近い改善が見られました。また、一次精母細胞で精子発生が停止している無精子症マウス2系統からもそれぞれ産子を得ることに成功しました(図3)。
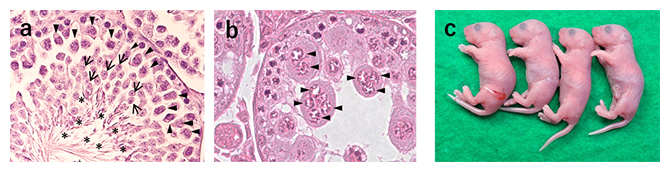
図3 無精子症マウスからの産子作出
コントロール(a)および無精子症マウス(Stx2repro34マウス)の精巣像(b)。Stx2repro34マウスでは一次精母細胞が合胞体化し、それ以降の精細胞が存在しない。a、bの記号はそれぞれ一次精母細胞(▲)、精子細胞(→)、精子(*)を示す。cはStx2repro34マウス由来一次精母細胞の顕微授精により得られた産子。
本研究で開発された顕微授精法によって得られた産子のうち、里親に哺育させた11匹は全て正常に発育し、子孫も作りました。また、マルチカラーFISH解析[8]を用いて全ての染色体を検査したところ、4匹で性染色体[9]の異常が見つかりました。この原因が、染色体異常が性染色体で生じやすいことによるのか、それとも常染色体[9]でも異常が生じているのかを明らかにするため、移植前の胚をマルチカラーFISH解析しました。その結果、常染色体・性染色体にかかわらず異常が生じていることが分かりました。このことより、正常な常染色体を所持した胚のみが選抜されて産子まで発生し、産まれた産子の一部に性染色体異常のみが残ることが明らかになりました。
今後の期待
本研究によって、一次精母細胞を用いた顕微授精が、用いる卵子の細胞質サイズを小さくすることによって、従来の精子や精子細胞を用いた顕微授精と同じように、実用的な技術になる可能性を示しました。現在、ヒト男性不妊のうち顕微授精で治療できるのは、減数分裂以降の精子・精子細胞がある場合に限定されています。一次精母細胞は多くの無精子症男性にもその存在が認められているので、将来、本技術が新たな不妊治療法の一つとなるかもしれません。
しかし、本技術がヒトで応用されるためには、染色体異常を改善するという大きなハードルがあります。また、卵子の細胞質サイズを小さくするだけでは染色体の異常の全てが修正されないことも分かっています。今後の研究により、なぜ一次精母細胞由来の染色体に異常が出やすいのか、どのような一次精母細胞の減数分裂異常が修正可能なのかなど、多くの疑問が解き明かされることが期待できます。
補足説明
1.一次精母細胞
精子の発生過程において、精祖(精原)細胞が分化して生じる精細胞。2回の減数分裂を経て最終的に精子となる。
2.顕微授精
顕微鏡下で、細いガラス針で精子を卵子に注入する受精補助技術。運動能を持たない精子や、精子細胞などの未成熟精子を用いた受精も可能にする。
3.減数分裂
真核生物の生殖細胞で見られる細胞分裂様式。2回の分裂を経て染色体数を半分に減らすことで、配偶子(精子と卵子)を作る。
4.無精子症
雄の個体において、精液中に精子が存在しない症状。ヒトでは男性の1%程度に認められる。
5.生殖補助技術
体外受精をはじめ、顕微授精、精子・卵子あるいは受精卵の凍結保存、融解移植など、受精から妊娠に関わる事象を人為的に手助けする技術。
6.染色体異常
染色体の構造的あるいは数的な異常。胚発生の停止や流産の原因となる。出産までたどり着いた場合でも、ヒトにおいてはダウン症やターナー症候群などの疾患の原因となる。
7.姉妹染色分体の早期分離
細胞が分裂の前にDNA複製を行い、出来上がった二つの同一な染色体を姉妹染色分体という。姉妹染色分体は、減数分裂の第二分裂期で分離するが、それよりも早い段階で分離が起こると、染色体数の異常が生じ、胚の発生停止や流産、ダウン症などの原因につながる。
8.マルチカラーFISH解析
個々の染色体に特異的に結合する蛍光プローブを用い、全染色体を別々の色に染色することで、各染色体の構造異常や異数性を解析する技術。
9.性染色体、常染色体
遺伝子で雌雄が決まる生物は、相同の2組の染色体(常染色体)と、性別を決定する遺伝子を含む染色体(性染色体)を持つ。哺乳類の性染色体はX染色体とY染色体から成る。例えばマウスの染色体は40本で、そのうち38本は常染色体、残りの2本は性染色体(雌のXXまたは雄のXY)で構成されている。
共同研究グループ
理化学研究所
バイオリソース研究センター 遺伝工学基盤技術室
専任技師 越後貫 成美(おごぬき なるみ)
専任研究員 井上 貴美子(いのうえ きみこ)
室長 小倉 淳郎(おぐら あつお)
(開拓研究本部 小倉発生遺伝工学研究室 主任研究員)
生命機能科学研究センター 染色体分配研究チーム
客員研究員 京極 博久(きょうごく ひろひさ)
(神戸大学 農学研究科 生殖生物学研究室 助教)
チームリーダー 北島 智也(きたじま ともや)
旭川医科大学 医学部 生物学教室
助教 日野 敏昭(ひの としあき)
教授 立野 裕幸(たての ひろゆき)
筑波大学 実験動物学研究室
大学院生(研究当時) 大澤 優生(おおさわ ゆき)
(現 日本新薬株式会社)
准教授 水野 聖哉(みずの せいや)
教授 杉山 文博(すぎやま ふみひろ)
東京大学 定量生命研究所 病理発生制御研究分野
助教 藤原 靖浩(ふじわら やすひろ)
岡山理科大学 獣医学部 獣医学科 獣医動物衛生学講座
教授 国枝 哲夫(くにえだ てつお)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業基盤研究(C)「ライブセルイメージング技術を用いた一次精母細胞顕微授精技術の改善(研究代表者:越後貫成美)」、同新学術領域研究「全能性プログラム:デコーディングからデザインへ(領域代表者および研究代表者:小倉淳郎、研究分担者:日野敏昭、研究代表者:京極博久)」「染色体イメージングによる卵子インテグリティの予見(研究代表者:北島智也)」による支援を受けて行われました。
原論文情報
Narumi Ogonuki, Hirohisa Kyogoku, Toshiaki Hino, Yuki Osawa, Yasuhiro Fujiwara, Kimiko Inoue, Tetsuo Kunieda, Seiya Mizuno, Hiroyuki Tateno, Fumihiro Sugiyama, Tomoya S. Kitajima, and Atsuo Ogura, “Birth of mice from meiotically arrested spermatocytes following biparental meiosis in halved oocytes”, EMBO Reprots, 10.15252/embr.202254992
発表者
理化学研究所
バイオリソース研究センター 遺伝工学基盤技術室
専任技師 越後貫 成美(おごぬき なるみ)
室長 小倉 淳郎(おぐら あつお)
(開拓研究本部 小倉発生遺伝工学研究室 主任研究員)
生命機能科学研究センター 染色体分配研究チーム
客員研究員 京極 博久(きょうごく ひろひさ)
チームリーダー 北島 智也(きたじま ともや)
報道担当
理化学研究所 広報室 報道担当

")