
2023-10-17 理化学研究所,東京薬科大学,横浜市立大学
理化学研究所(理研)生命機能科学研究センター エピジェネティクス制御研究チーム(研究当時)の梅原 崇史 チームリーダー(研究当時、現 創薬タンパク質解析基盤ユニット 上級研究員)、菊地 正樹 研究員(研究当時、現 生命医科学研究センター 免疫器官形成研究チーム 研究員)、環境資源科学研究センター ケミカルゲノミクス研究グループの伊藤 昭博 客員主管研究員(東京薬科大学 生命科学部 教授)、横浜市立大学大学院 生命医科学研究科 構造エピゲノム科学研究室の小沼 剛 助教らの共同研究グループは、多くのタイプのがん細胞で高発現しているタンパク質GAS41[1]が、後成遺伝情報[2](エピジェネティック情報)を担うヒストンH3タンパク質のアセチル化修飾[3]を認識し、特定の遺伝子の発現を活性化する仕組みを発見しました。
本研究成果により、GAS41が高発現している多種類のがん細胞の増殖を抑える制御分子の合理的な開発が期待されます。
ヒトをはじめとする真核細胞[4]のゲノムDNAはヌクレオソーム[5]という構造を形成して凝縮しており、エピゲノム[2]はどの遺伝子のヌクレオソームをその転写[6]に先立って解きほぐすかを制御しています。
今回、共同研究グループは構造生物学・生化学・細胞生物学の手法でGAS41の構造と機能を解析しました。その結果、GAS41タンパク質が、ヒストンH3の「N末端[7]」と「N末端から数えて14番目または27番目のリシン残基[8]のアセチル化修飾」をそれぞれ異なるポケットを用いて認識し、ヌクレオソームにH2A.Zというヒストンの変種タンパク質の導入を促進することで、特定の遺伝子を転写しやすくする仕組みが明らかになりました。
本研究は、科学雑誌『Proceedings of the National Academy of Sciences(PNAS)』オンライン版(10月16日付)に掲載されました。
背景
後成遺伝情報はエピゲノムから転写する遺伝子の質と量の制御に関わることが知られています。数多く知られている後成遺伝情報の中でもヒストンのアセチル化修飾は、多細胞生物のそれぞれの細胞種に特異的に発現している遺伝子の転写を活性化する上で重要と考えられています。近年、ヌクレオソーム内部のヒストンにおけるリシン残基のアセチル化がどのような順序で伝播して遺伝子の転写を活性化するかの仕組みや、ヒストンの特定のリシン残基のアセチル化がエピゲノムからの遺伝子転写のどの反応素過程を活性化するかの仕組みなどが示唆されています注1、2)。
ヒストンのアセチル化修飾が遺伝子転写の活性化を引き起こす仕組みには複数の経路があります。重要な制御経路として、アセチル化されたヒストンの種類やリシン残基の位置を見分けて認識する「読み手」タンパク質が関わる経路が知られています。ヒストンのアセチル化修飾に対しては、ブロモドメイン[9]やYEATSドメイン[1]を持つタンパク質が主要な「読み手」タンパク質であり、細胞内でこれらのタンパク質の発現が亢進するとがんや炎症などの疾患につながることが知られてきています。
ブロモドメインがヒストンのアセチル化修飾を認識する仕組みについては解析が進んでおり、特定のブロモドメインを持つタンパク質の機能を選択的に阻害する化合物の開発やそれを利用したがん制御機構の研究が進められてきています注3、4)。その一方、YEATSドメインがヒストンのアセチル化修飾を認識する仕組みについては不明な点が多く、YEATSドメインに対する阻害剤の開発も遅れている状況でした。
そこで共同研究グループは、ヒトに4種類存在するYEATSドメイン含有タンパク質のうち、難治性がんの一種である膠芽腫(こうがしゅ)細胞[10]で遺伝子増幅が見られるGAS41に着目しました。GAS41は、真核生物の種間で保存されたクロマチン[5]の構造を制御するタンパク質複合体の構成成分の一つであり、クロマチンの構造を変化させることで遺伝子の転写を活性化します。GAS41の遺伝子は膠芽腫細胞だけでなく、胃がんや肝がん、直腸がん、すい臓がん、乳がんや肺がんの細胞で過剰発現が見られ、細胞内でGAS41遺伝子の発現を弱めるとこのような発がんを軽減できることが近年知られています。GAS41による発がん活性は、GAS41のYEATSドメインがヒストンのアセチル化修飾を認識することと関連していますが、その認識がどのような仕組みによって担われているのかについては不明な点がありました。そこで本研究では、GAS41がYEATSドメインを介してどのようにヒストンの特定のアセチル化状態を認識して細胞内で機能するのかの仕組みの解明を目指しました。
注1)2023年7月17日プレスリリース「遺伝子発現を制御するエピゲノムの複製と転写」
注2)2020年11月26日プレスリリース「エピゲノムの制御を受けた転写の方程式」
注3)2018年8月8日プレスリリース「『がんエピゲノム』を検出する新手法」
注4)2023年10月3日プレスリリース「遺伝子発現を活性化するスーパーエンハンサーの再定義」
研究手法と成果
共同研究グループは初めに、GAS41のYEATSドメインがヒストンのどのようなアセチル化状態を認識するのかについて、ヒストンの種類、N末端テイルにおけるアセチル化修飾の位置、N末端テイルの長さを変えて生化学的に比較検討しました。その結果、GAS41のYEATSドメインはヒストンH3のN末端から数えて14番目のリシン残基のアセチル化(K14ac)と最も強く結合すること、さらにその結合にはヒストンH3のN末端1~6番目までのアミノ酸残基が不可欠なことが分かりました。通常、「読み手」タンパク質は、特定の化学修飾を含むアミノ酸残基とその前後の数カ所の残基のアミノ酸配列を認識します。今回のデータは、GAS41のYEATSドメインがヒストンH3の「N末端」と「K14ac」という離れた2カ所を何らかの方法で認識する仕組みを備えていることを示唆しました。
そこで次に、K14acを含むヒストンH3のN末端テイル領域とGAS41のYEATSドメインとの複合体の結晶をX線で構造解析しました。その結果、リシンアセチル化を「読む」機能を持つGAS41のYEATSドメインは確かにアセチル化したヒストンH3テイルのK14acとその周辺のアミノ酸配列に結合していました(図1、左端のパネル)。この結合は、YEATSドメインが形成する芳香族ケージと呼ばれるポケットによって担われています。

図1 GAS41がヒストンH3のN末端テイルを認識する2種類の仕組み
左側のパネル:左側3枚のパネルのうちの中央のパネルがGAS41のYEATSドメインとヒストンH3ペプチドとの複合体の結晶構造の全体を示し、その左右のパネルに2箇所の結合場所の拡大図を示している。ヒストンH3のペプチドを紫色のスティックで、GAS41のYEATSドメインをベージュでそれぞれ示す。各残基の位置は、アミノ酸の種類(1文字表記)と残基の位置(数字)のラベルで示す。
右端のパネル:GAS41のYEATSドメインの分子表面モデル。分子表面の疎水性の度合いを緑色から白色のグラデーションで示す。より白色の分子表面がより疎水的である。ヒストンH3のN末端テイルの1番目のアラニン(A1)から3番目のスレオニン(T3)までのアミノ酸残基がGAS41のYEATSドメインのくぼみ(H3NTポケット)によって認識されている。
さらにGAS41のYEATSドメインは、この芳香族ケージによる結合部位から離れた場所で、上記とは別のヒストンH3テイルのN末端領域と結合することが分かりました(図1、左から2番目と3番目のパネル)。この第2の結合場所では、YEATSドメインの疎水的な分子表面のくぼみ(H3NTポケットと命名)がH3テイルのN末端1~3番目までの3残基を認識していることが分かりました(図1、右端のパネル)。
X線結晶構造解析では、タンパク質を結晶化(固体化)して解析するため、得られた構造が生理的な状況を反映していない可能性があります。今回の解析でも、一つのGAS41タンパク質に二つのヒストンH3タンパク質が結合している構造が得られており、これが実際に水溶液中や細胞内の姿であるかを検証する必要がありました。そこで次に、NMR法[11]によりGAS41のYEATSドメインが水溶液中でどのようにヒストンH3テイルと相互作用するのかを調べました。その結果、ヒストンH3テイルの14番目のリシン(K14ac)や27番目のリシン(K27ac)がアセチル化されていると、GAS41のYEATSドメインは芳香族ケージとH3NTポケットの間に位置するアミノ酸(115番目のチロシン残基:Y115)を介してH3ペプチドと相互作用することが分かりました(図2)。
この結果から、GAS41のYEATSドメインは、水溶液中では1本のH3テイルの2カ所の異なる部位を2カ所のポケットを介して同時に認識する2価認識モデルが示唆されました。また、H3NTポケットを変異させたタンパク質を培養細胞で発現させる実験を行った結果からも、結晶構造で観察されたGAS41のH3NTポケットとヒストンH3のN末端との相互作用が細胞内でも起きていることが示唆されました(図3A)。
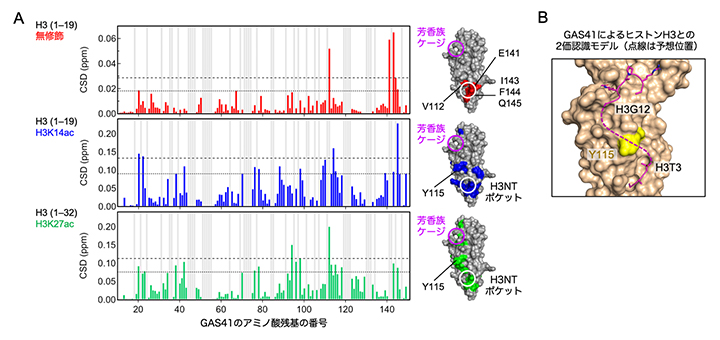
図2 水溶液中でGAS41がヒストンH3のN末端テイルと相互作用する部位
(A)溶液NMRによるGAS41のYEATSドメインの残基の化学シフトの比較。無修飾H3ペプチド(上)、K14アセチル化H3ペプチド(中)、K27アセチル化H3ペプチド(下)の滴定前後でのシグナルの変化を示す。Y軸のCSDは化学シフトの差分を示す。CSDが大きく変化しているアミノ酸残基は、滴定したそれぞれのH3ペプチドとの相互作用が強いことを示している。X軸に平行な2種類の破線は相互作用の有意差を検定する基準であり、上が平均値に2標準偏差、下が平均値に1標準偏差を加えた値を示す。各データの右側にそれぞれのペプチドと有意に相互作用した残基をGAS41のYEATSドメインの表面に色でマッピングし、残基番号をラベルで示した。芳香族ケージの位置は紫色の丸印で、H3NTポケットの位置は白色の丸印で示す。
(B)GAS41のYEATSドメイン表面の構造における115番目のチロシン残基の位置(黄色)。2本のK14アセチル化H3ペプチドの実際の構造(紫色の実線)は模式的に点線で結ばれている。芳香族ケージに結合するH3ペプチドの12番目のグリシン残基(H3G12)と、H3NTポケットに結合するH3ペプチドの3番目のスレオニン残基(H3T3)の位置をラベルで示す。Aの解析において、H3ペプチドがアセチル化されていると、両方の残基の間に位置する115番目のチロシン残基(Y115)に有意な化学シフト変化が検出されたことから、水溶液中ではGAS41のYEATSドメインは1本のH3テイルを2カ所の異なるポケットで(=2価で)認識することが示唆された。
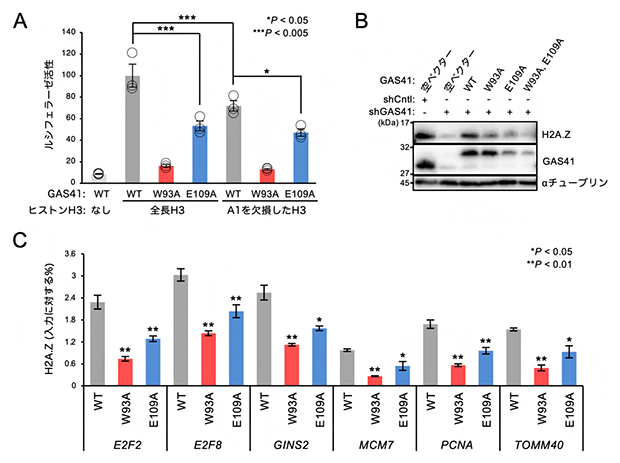
図3 GAS41によるヒストンH3のN末端認識は細胞内機能に重要
(A)GAS41とヒストンH3との細胞内での結合評価。細胞内でGAS41の野生型(WT)または2種類の機能変異体(W93AまたはE109A)と、全長のヒストンH3またはN末端認識の意義を調べるために1番目のアラニン残基(A1)を欠損したヒストンH3を発現させ、細胞内で2種類のタンパク質間の相互作用を検出する測定方法であるNanoBiTアッセイで蛍光強度(Y軸のルシフェラーゼ活性)を測定した。W93AはYEATSドメインの芳香族ケージの機能変異体、E109AはYEATSドメインのH3NTポケットの機能変異体のGAS41をそれぞれ示す。データは3回の独立実験の平均±標準誤差。Holm検定による統計的有意性を*または***で示す。
(B)野生型または変異体のGAS41を導入したGAS41発現抑制細胞におけるGAS41とH2A.Zタンパク質の免疫ブロット解析。shCntlは対照実験、shGAS41はGAS41を発現抑制した実験(+は処理、-は未処理)を示す。機能変異体においてはGAS41のプロットの濃さに対するH2A.Zのプロットの濃さの比が低くなっており、H2A.Zタンパク質の量が減少していることが分かる。
(C)野生型または変異型のGAS41を導入したGAS41発現抑制細胞におけるGAS41標的遺伝子プロモーターでのクロマチン免疫沈降-定量リアルタイムPCR(ChIP-qPCR)の解析結果。クロマチン免疫沈降(ChIP)は、細胞のクロマチンを断片化し、特定の抗体が認識するタンパク質やその化学修飾の状態を含むクロマチン断片を取得する方法。定量リアルタイムPCR(qPCR)は、測定する試料に含まれるDNAの量の推定が可能なDNAの増幅法。この2種類の手法を組み合わせることで、クロマチンの特定の領域に含まれているタンパク質やその修飾状態の量(クロマチンにおける占有率)を測定できる。このChIP-qPCR解析では、ヒストンH2A.Zタンパク質の占有率を抗体で検出した。各GAS41標的遺伝子を下部に示した。データは3回の独立実験の平均±標準偏差。Welchの両側t検定による統計的有意性を*または**で示す。
GAS41の機能として、ヌクレオソームにヒストンの変種タンパク質であるH2A.Zタンパク質の導入を促進することで、特定の遺伝子を転写しやすくすることが知られています。そこで次に、GAS41とヒストンH3との相互作用がGAS41の細胞機能に必要かどうかを確認するために、GAS41によるH2A.Zタンパク質の量的な制御に対する影響を調べました。GAS41の遺伝子発現抑制によってH2A.Zの量が減少している細胞を用いて、野生型GAS41、H3NTポケットあるいは芳香族ケージを変異させたGAS41を発現させた結果、変異体を発現した細胞ではH2A.Zタンパク質の量が減少することが分かりました(図3B)。従って、GAS41のH3NTポケットと芳香族ケージは、細胞内でH2A.Zタンパク質の量を維持するために重要なことが分かりました。
GAS41はクロマチンの構造を制御するタンパク質複合体の構成成分の一つであり、特定の遺伝子プロモーター[12]領域のヌクレオソームにH2A.Zの導入を促進することでクロマチンの構造を変化させ、その遺伝子の転写を活性化します。そこで最後に、GAS41が標的としている遺伝子プロモーター上のH2A.Zのクロマチンにおける占有率を調べました(図3C)。これまで知られている6種類の遺伝子プロモーターをGAS41の標的として選択し、GAS41の野生型、またはH3NTポケットあるいは芳香族ケージの変異体を細胞に発現してこれらの遺伝子プロモーターに結合するヒストンH2A.Zの量を検討しました。その結果、これら6種類の遺伝子プロモーターのすべてにおいて、それぞれの変異体を発現する細胞ではH2A.Zのクロマチンにおける占有率が有意に低下しました。これらの結果から、GAS41は芳香族ケージを介してヒストンH3のアセチル化状態を認識するだけでなく、H3NTポケットを介してヒストンH3のN末端を認識することにより、標的とする遺伝子のプロモーターを精密に認識し、その遺伝子の転写の活性化に寄与する仕組みが示唆されました(図4)。
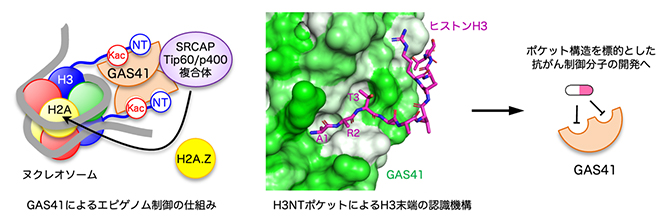
図4 GAS41によるエピゲノム制御の仕組みと創薬への応用
左図は、GAS41によるエピゲノム制御の仕組みを模式的に示す。GAS41はSRCAPまたはTip60/p400と呼ばれるタンパク質複合体を構成するタンパク質であり、ヌクレオソームを構成するヒストンH3のN末端テイルのN末端(NTで示す)とK14acまたはK27ac(Kacで示す)を同時に認識する。これにより、GAS41の2量体を含むSRCAP複合体またはTip60/p400複合体は特定の遺伝子プロモーターのヌクレオソームのアセチル化状態を正しく認識し、そのヌクレオソームにヒストンH2A.Zを促進的に導入することで特定の遺伝子の転写開始反応を活性化すると考えられる。右図は、今回見いだしたGAS41のH3NTポケットの構造情報が抗がん制御分子の合理的な開発に資するイメージを示す。
今後の期待
多くのがん細胞や炎症細胞などでヒストンのアセチル化状態の異常が見られることから、ヒストン脱アセチル化酵素に対する阻害剤は難治性のがんに対する治療薬として実用化されています。ヒストンのアセチル化認識を阻害する化合物についても、ブロモドメインに対する阻害剤は親和性や選択性に優れた化合物が多数開発され、数多くのがん種に対する臨床試験が進行中です。その一方、YEATSドメインに対する阻害剤はブロモドメイン阻害剤ほど開発が進んでおらず、特にGAS41に対する阻害剤の開発は遅れている状況です。
今回の研究により、GAS41のYEATSドメインには、他のタンパク質のYEATSドメインには見られない機能的なポケットが存在することが分かりました。GAS41の発現は膠芽腫細胞だけでなく、胃がん細胞や肺がん細胞などにおいても異常があることが知られています。今後、今回明らかになった構造情報を活用することで、がん制御に向けてGAS41を選択的に阻害する化合物を合理的に開発することが期待されます。
補足説明
1.GAS41、YEATSドメイン
GAS41(膠芽腫(こうがしゅ)増幅配列41)は、ヒストンH3のN末端テイルの化学修飾を認識するタンパク質であり、ヒトでは全長が227アミノ酸から成る。分子内にYEATSドメインとコイルド・コイルドメインと呼ばれる2種類のドメインを持つ。YEATSドメインは120~140アミノ酸で構成される構造を持つドメインであり、ヒストンのリシン残基のアセチル化修飾やアシル化修飾を認識する役割を持つ。コイルド・コイルドメインはコイルド・コイルドメイン同士での相互作用を介してタンパク質の2量体を形成する役割を持つ。
2.後成遺伝情報、エピゲノム
細胞内のDNAには遺伝情報が塩基配列として記録されている。細胞には、DNAの塩基配列以外にも細胞の個性を記憶する個々の情報が存在しており、それらはDNAやヒストンなどに対する化学修飾として記録されている。このDNAの周辺に記録されるDNAの塩基配列以外の生命情報を後成遺伝情報(エピジェネティック情報)と呼ぶ。「ゲノム」が細胞内の全てのDNAの塩基配列として記録された遺伝情報の総体を指すのに対し、「エピゲノム」はDNAやヒストンなどの化学修飾によって細胞の個性を記憶する情報を含めた総体を指す。
3.ヒストンH3タンパク質のアセチル化修飾
ヒストンと呼ばれるタンパク質はDNAを巻き付けることで長大なDNAを核内に納める役割を持つ。ヒストンはDNAを巻き付けるために、ヒストンH3とH4が2個ずつ集まったH3-H4四量体一つとヒストンH2AとH2Bが1個ずつ集まったH2A-H2B二量体二つから成るヒストン八量体を形成している。ヒストンは生物種間を超えて高く保存されており、特定のアミノ酸残基が化学修飾されている例が数多く知られている。代表的な化学修飾であるアセチル化修飾の場合、タンパク質のリシン残基の側鎖アミンにアセチル基(CH3CO-)が結合する。ヒストンH3の場合、N末端側の1残基目から数えて4番目、9番目、14番目、18番目、23番目、27番目、36番目などのリシン残基がアセチル化酵素によってアセチル化され、そのうちの多くは遺伝子転写の活性化状態と相関している。
4.真核細胞
真核細胞は核膜で包まれた核やその他の小器官を持つ細胞のこと。真核細胞のゲノムDNAは、ヌクレオソームを基本単位とする凝縮した構造を形成して、細胞周期の大部分を占める間期においては細胞核の中に収納された状態で存在する。真核細胞で構成される真核生物の代表的な生物としては単細胞で構成される酵母や、多細胞で構成される昆虫やマウス、ヒトなどが挙げられる。
5.ヌクレオソーム、クロマチン
真核細胞核内でDNAとヒストン八量体(H2A、H2B、H3、H4のタンパク質を2個ずつ含む複合体)が周期的に巻き付いて形成された複合体をヌクレオソームと呼ぶ。ヌクレオソームの中心部分は145~147塩基対のDNAがヒストン八量体に巻き付いたコア粒子から成る。長鎖のDNAと複数のヌクレオソームで構成される数珠状の構造体はクロマチンと呼ばれる。
6.転写
遺伝子DNAの塩基配列をRNAポリメラーゼが読み取り、2本鎖DNAの片方の塩基配列に対応するRNAを合成する反応を「転写」と呼ぶ。RNAポリメラーゼは、DNAを鋳型としてRNAを構成する基本単位のリボヌクレオチドを重合する酵素であり、原核生物では1種類のRNAポリメラーゼ、真核生物ではRNAポリメラーゼⅠ、Ⅱ、Ⅲの3種類が存在する。多くの遺伝子はその本体部分だけでは転写が起こらずに、遺伝子の上流または下流に転写反応を質的・量的に変化させるDNA配列が必要である。これらはその位置や機能に応じてプロモーターやエンハンサーと呼ばれる。
7.N末端
タンパク質はアミノ酸残基が直鎖状に連結したポリペプチドで構成されることから、その始まりと終わりがある。タンパク質の始まりとなる1番目の残基(またはその主鎖アミノ基)がN末端、終わりとなる最後の残基(またはその主鎖カルボキシル基)がC末端と呼ばれる。タンパク質によってはN末端残基のアミノ基は細胞内でアセチル化修飾を受けているが、ヒストンH3のN末端残基のアミノ基は化学修飾を受けていない。また、ヒストンのN末端残基に続く数十残基の領域は特定の構造を持たないテイル(尾部)として水溶液中で揺らいでいる。ヒストンN末端テイルのリシン残基側鎖のアミノ基は、アセチル化やメチル化などの化学修飾を受けやすく、その特定の修飾状態は遺伝子発現の活性化や抑制化の状態と相関している。
8.リシン残基
タンパク質を構成するアミノ酸の一つ。1文字略称はK。側鎖にアミノ基を持つため、タンパク質中の残基としてアミノ基へのアセチル化やメチル化修飾の標的となる。
9.ブロモドメイン
110~120アミノ酸から成るタンパク質の折り畳み構造。この折り畳み構造の中心部分にくぼみを持ち、このくぼみを介してヒストンのN末端テイルなどに存在するリシン残基のアセチル化状態を特異的に認識できる。
10.膠芽腫(こうがしゅ)細胞
脳内に生じる悪性度が高い腫瘍の一種。
11.NMR法
分子(タンパク質など)に対して高磁場の環境下で電磁波を照射し、分子を構成する原子のそれぞれ原子核が周辺の化学的環境に応じて特定の電磁波を吸収する共鳴現象を観測することで分子の立体構造やその変化を推定する手法。原子核の種類が同じでも原子核の周辺環境の違いによって共鳴周波数は変化し、化学シフトと呼ばれる分子の立体構造に関する情報が得られる。
12.遺伝子プロモーター
DNA上で遺伝子(RNA)の転写が開始される位置の近くにあり、遺伝子を発現させる機能を持つ塩基配列。遺伝子プロモーターに数多くの基本転写因子とRNAポリメラーゼが結合することで、特定の遺伝子が転写される位置や転写される量が決まると考えられている。
共同研究グループ
理化学研究所
生命機能科学研究センター
エピジェネティクス制御研究チーム(研究当時)
チームリーダー(研究当時)梅原 崇史(ウメハラ・タカシ)
(現 創薬タンパク質解析基盤ユニット 上級研究員)
研究員(研究当時)菊地 正樹(キクチ・マサキ)
(現 生命医科学研究センター 免疫器官形成研究チーム 研究員)
環境資源科学研究センター
ケミカルゲノミクス研究グループ
客員主管研究員 伊藤 昭博(イトウ・アキヒロ)
(東京薬科大学 生命科学部 教授)
東京薬科大学
生命科学部 細胞情報科学研究室
特別研究員(PD)(研究当時)高瀬 翔平(タカセ・ショウヘイ)
研究員 則次 恒太(ノリツグ・コウタ)
修士課程2年生(研究当時)関根 咲彩(セキネ・サアヤ)
横浜市立大学
大学院生命医科学研究科 構造エピゲノム科学研究室
助教 小沼 剛(コヌマ・ツヨシ)
教授 池上 貴久(イケガミ・タカヒサ)
研究支援
本研究は、理化学研究所運営費交付金(生命機能科学研究)で実施し、科学技術振興機構(JST)戦略的創造研究推進事業個人型研究(PRESTO、さきがけ)「細胞機能の構成的な理解と制御(研究領域総括:上田泰己)」の研究課題「『エピヌクレオソーム』の精密な再構成による遺伝子発現制御解析(研究代表者:梅原崇史)」、日本学術振興会(JSPS)科学研究費助成事業学術変革領域研究(A)「DNAの物性から理解するゲノムモダリティ(領域代表者:西山朋子)」、同基盤研究(B)「がん細胞で頑強に維持される超アセチル化エピゲノムを操作する(研究代表者:梅原崇史)」、同若手研究「GAS41によるアセチル化ヌクレオソーム認識機構の解明(研究代表者:菊地正樹)」などによる助成を受けて行われました。
原論文情報
Masaki Kikuchi, Shohei Takase, Tsuyoshi Konuma, Kota Noritsugu, Saaya Sekine, Takahisa Ikegami, Akihiro Ito, and Takashi Umehara*, “GAS41 promotes H2A.Z deposition through recognition of the N terminus of histone H3 by the YEATS domain”, Proceedings of the National Academy of Sciences of the United States of America(PNAS), 10.1073/pnas.2304103120
発表者
理化学研究所
生命機能科学研究センター エピジェネティクス制御研究チーム(研究当時)
チームリーダー(研究当時)梅原 崇史(ウメハラ・タカシ)
(現 創薬タンパク質解析基盤ユニット 上級研究員)
研究員(研究当時)菊地 正樹(キクチ・マサキ)
(現 生命医科学研究センター 免疫器官形成研究チーム 研究員)
環境資源科学研究センター ケミカルゲノミクス研究グループ
客員主管研究員 伊藤 昭博(イトウ・アキヒロ)
(東京薬科大学 生命科学部 教授)
横浜市立大学
大学院生命医科学研究科 構造エピゲノム科学研究室
助教 小沼 剛(コヌマ・ツヨシ)
報道担当
理化学研究所 広報室 報道担当
東京薬科大学 総務部 広報課
横浜市立大学 広報課長 上村一太郎

