2018-05-11 理化学研究所
理化学研究所(理研)生命機能科学研究センター染色体分配チームのイ・ディン リサーチアソシエイトと北島智也チームリーダーらの国際共同研究チーム※は、正しい染色体数を持つ卵子を生むために減数分裂[1]の過程で働く新たな分子経路を発見しました(図)。
本研究成果は、先天性疾患の原因となる卵子の染色体数異常[2]の解明に貢献すると期待できます。
正常な染色体数を持つ卵子は、前駆細胞である卵母細胞の減数分裂によって生まれます。卵母細胞から卵子に染色体が正しく分配されるためには、複製された姉妹染色分体[3]ペアをつなぎ留めているセントロメア[4]接着が適切に維持されることが重要です。タンパク質シュゴシン[5]は、減数分裂のある時期までセントロメア接着を保護していることが分かっています。しかし、シュゴシンが見られなくなった後のセントロメアで、どのように接着が維持されているのかは明らかになっていませんでした。今回研究チームは、マウスの卵母細胞を用いた研究から、シュゴシンが観察されなくなった後には、「SUMO[6]化」と呼ばれるタンパク質修飾経路が強化されることで、セントロメア接着が維持されることを見いだしました。セントロメア接着の維持の異常は、ヒト卵子で染色体数異常が生じる原因の一つであると考えられ、今回の発見は、卵子における染色体数異常の原因の分子的な理解につながる可能性があります。
本研究は、米国の科学雑誌『Current Biology』(5月21日号)の掲載に先立ち、オンライン版(5月10日付け:日本時間5月11日)に掲載されます。
※国際共同研究チーム
理化学研究所
生命機能科学研究センター 染色体分配研究チーム
チームリーダー 北島 智也(きたじま ともや)
(旧 多細胞システム形成研究センター 染色体分配研究チーム)
リサーチアソシエイト イ・ディン(Yi Ding)
(旧 多細胞システム形成研究センター 染色体分配研究チーム)
多細胞システム形成研究センター 染色体分配研究チーム(研究当時)
テクニカルスタッフ 海道 雅子(かいどう まさこ)
スペイン国立研究評議会-サラマンカ大学
教授 アルベルト・ペンダス(Alberto M. Pendas)
教授 エレナ・ルラノ(Elena Llano)
※研究支援
本研究は、日本学術振興会(JSPS)科学研究費補助金若手研究A「卵母細胞に特有な動原体の役割(研究代表者:北島智也)」等の支援を受けて行われました。
背景
卵子の染色体数異常は、流産やダウン症[7]などの先天性疾患の主要な原因です。卵子の染色体数は、卵子の前駆細胞である卵母細胞の減数分裂における染色体分配によって決定されます。ここで重要な役割を果たすのが、複製された姉妹染色分体ペアをつなぎ留めているセントロメア接着です(図1)。減数分裂は2回の連続した細胞分裂から成り、相同染色体[8]を分配する減数第一分裂では、セントロメア接着は維持されたままです。次の減数第二分裂でセントロメア接着が解除されることで、姉妹染色分体が正しく分配されます。
減数第一分裂でセントロメア接着を維持する機構の一つとして知られているのが、セントロメアに局在するタンパク質シュゴシンによる接着の保護です。減数第一分裂の後期が開始すると、相同染色体の分配のために染色体腕部における姉妹染色分体ペア間の接着が解除される一方、セントロメアにおいてはシュゴシンによって接着が保護されることで、姉妹染色分体はつなぎ留められたままになります。しかし、シュゴシンは減数第一分裂の後期開始を過ぎると、速やかにセントロメアから見られなくなってしまいます。シュゴシンが見られなくなった後のセントロメアにおいて、減数第一分裂の終了まで接着を維持する機構については、これまで探求されたことはほとんどありませんでした。
研究手法と成果
国際共同研究チームは、哺乳類卵母細胞のセントロメア接着を保護する未知の制御因子が存在すると考え、セントロメアに結合する性質を持つタンパク質を酵母2-ハイブリッド法[9]により探索しました。その結果、「SUMO化」と呼ばれるタンパク質修飾を行う酵素「PIAS1[10]」が同定されました。PIAS1はPIASファミリータンパク質の一つで、SUMOという小さなタンパク質をさまざまなタンパク質に結合させ、そのタンパク質の機能を制御する機能があります。
そこで、SUMO化に関わるタンパク質の局在を調べたところ、4種類のPIASタンパク質(PIAS1/2/3/4)と2種類のSUMOタンパク質(SUMO2/3)が、卵母細胞のセントロメアに局在することが分かりました。興味深いことに、これらの局在は減数分裂の進行とともに変化し、しかもシュゴシンとは対照的な傾向が見られました。すなわち、シュゴシンは減数第一分裂後期開始を過ぎると速やかにセントロメアから見えなくなるのに対し、PIASやSUMOのセントロメア局在は、減数第一分裂後期から終期にかけて増えていました(図2)。
次に、セントロメアにおけるSUMO化の役割について調べました。セントロメアに局在するPIASタンパク質の酵素活性を阻害し、SUMO化の機能を失わせた卵母細胞を観察したところ、減数第一分裂の後期開始においてシュゴシンによるセントロメア接着の保護に異常は見られませんでした。しかし、続いて起こる終期において、セントロメア接着が失われてしまうことが分かりました。さらに、セントロメアSUMO化の機能を後期開始よりも後の時期で回復させると、減数第一分裂を通してセントロメア接着が維持されるようになりました。これらの結果から、セントロメアSUMO化は減数第一分裂の後期開始よりも後の時期において、セントロメア接着に必要であることが示されました(図3)。
以上の結果から、セントロメアSUMO化は卵子に正しい数の染色体を持たせるための、シュゴシンとは独立に働く新しい経路として位置づけられます。セントロメアSUMO化は、減数第一分裂の後期開始以降、シュゴシンが見られなくなったセントロメアでそれを補うかのように強化され、減数第一分裂の終了までセントロメア接着の維持を保証することが示されました(図4)。
今後の期待
ヒト卵母細胞においては、母体の年齢とともにセントロメア接着が失われた染色体の頻度が上昇し、卵子の染色体数異常が生じると考えられています。これまで不明だった減数第一分裂後期開始後のセントロメア接着の維持に関わる分子経路の発見は、卵子の老化と染色体数異常の関係を分子的に理解する手がかりとなると期待できます。
原論文情報
Yi Ding, Masako Kaido, Elena Llano, Alberto M. Pendas, and Tomoya S. Kitajima, “The post-anaphase SUMO pathway ensures the maintenance of centromeric cohesion through meiosis I-II transition in mammalian oocytes”, Current Biology, 10.1016/j.cub.2018.04.019
発表者
理化学研究所
生命機能科学研究センター 染色体分配研究チーム
チームリーダー 北島 智也(きたじま ともや)
リサーチアソシエイト イ・ディン(Yi Ding)
お問い合わせ先
理化学研究所 生命機能科学研究センター センター長室 報道担当
山岸 敦(やまぎし あつし)
報道担当
理化学研究所 広報室 報道担当
補足説明
-
- 減数分裂
- 真核生物の生殖器官(精巣や卵巣)にある生殖細胞で見られる、配偶子(精子や卵子)を作るための特別な様式の細胞分裂。染色体セットの数が半分に減少して配偶子にもたらされるために、減数分裂と呼ぶ。
-
- 染色体数異常
- 多くの真核生物は決まった数の染色体数を持ち、有性生殖を行う種では配偶子の染色体数(n)に対して体細胞の染色体数はその2倍(2n)となる。細胞がこれらとは異なる染色体数を持つ場合、染色体数異常または異数性と呼ぶ。流産胚やダウン症で見られる染色体数異常の主な原因は、卵母細胞の減数第一分裂における染色体分配の誤りである。
-
- 姉妹染色分体
- 細胞は分裂の前にDNA複製を行う。染色体は複製によりできた二つの同一なDNAを含み、これらを姉妹染色分体と呼ぶ。姉妹染色分体の分配は、体細胞分裂では分裂後期に、減数分裂では第二分裂後期に起きる。
-
- セントロメア
- 細胞分裂に伴って複製された同一の染色体(染色分体)をつなぎとめ、紡錘糸が結合する領域をセントロメアと呼ぶ。ヒトでは、セントロメアは染色体の中央付近に位置することが多いため、分裂期の染色体はX字型に見えるが、マウスでは染色体の末端にセントロメアがあり、分裂期の染色体はV字型に見える。
-
- シュゴシン
- 細胞分裂の過程で染色分体をつなぎとめているタンパク質コヒーシンは、染色体分配の際に酵素によって分解される。この分解が減数分裂の不適切な時期に起きないようにコヒーシンを守っているタンパク質が、2004年に北島智也(当時東京大学)らにより発見され、日本語の「守護神」からシュゴシン(Shugoshin)と名付けられた。
-
- SUMO
- SUMOはSmall Ubiquitin-related Modifierの略。ユビキチン様タンパク質とも呼ばれる低分子量タンパク質。ユビキチンと同様に連結酵素の働きによりさまざまなタンパク質と結合するが、ユビキチンのようにタンパク質分解の目印となるのではなく、ユビキチン化によるタンパク質分解からの保護や、タンパク質の機能修飾を行うと考えられている。
-
- ダウン症
- 21番染色体が1本余分に存在し3本となるために引き起こされる先天性疾患。約9割が卵母細胞の減数分裂における染色体分配の誤りに起因している。
-
- 相同染色体
- 母方の配偶子(卵子)と父方の配偶子(精子)を由来とし、同じ大きさ・形・遺伝子構成を持つ一組の染色体。ヒトの場合は、22組の常染色体と、XXの性染色体が相同の関係にある。
-
- 酵母2-ハイブリッド法
- 転写活性化因子が持つDNA結合能と転写活性化能の2つの機能を応用した、タンパク質の結合因子をスクリーニングする手法の一つ。DNA結合領域との融合タンパク質と、転写活性化領域との融合タンパク質の2種類の融合(hybrid)タンパク質を作製し、それらを酵母に同時に発現させた際のレポーター遺伝子の活性化を指標に2つの融合タンパク質間の相互作用を判定する。本実験では、DNA結合領域との融合タンパク質としてセントロメアに局在するタンパク質CENP-Cを用い、マウス卵母細胞由来のcDNAライブラリーを転写活性化領域との融合タンパク質として酵母に発現させ、CENP-Cと相互作用するタンパク質の探索を行った。
-
- PIAS1
- タンパク質のSUMO化修飾は、SUMO化酵素と、脱SUMO化酵素により制御される。PIASは標的タンパク質にSUMOを結合する酵素(SUMOリガーゼ)の一種。PIAS1はProtein Inhibitor of Activated STAT-1の略。

図1 セントロメア接着、姉妹染色分体、相同染色体の関係
1本の染色体は姉妹染色分体ペアからなり、姉妹染色分体のペア間は紡錘糸が結合する染色体領域(セントロメア)や腕部で接着する。ヒトの染色体では、セントロメアが中央付近に位置するものが多く、分裂期の染色体はX字型に見えるものが目立つ(左)。本研究で用いたマウスでは、染色体の末端にセントロメアがあり、分裂期の染色体はV字型に見える(右)。
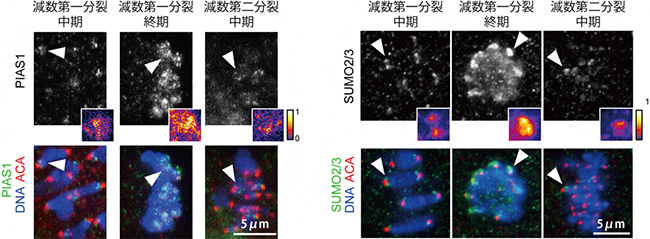
図2 SUMO化に関わるタンパク質の減数分裂期の局在
抗PIAS1抗体および抗SUMO2/3抗体を用いた蛍光免疫染色により、それぞれのタンパク質の局在を観察した。上段は、PIAS1もしくはSUMO2/3単独での蛍光シグナルを示す。小さな正方形は、白い矢頭で示した部分の拡大図で、シグナルの強さを炎色で表現したもの。黄色いほど局在が強いことを示している。下段は、PIAS1もしくはSUMO2/3(緑色)とDNA(青色)、セントロメア領域(ACA:赤色)を同時に観察した蛍光画像。PIAS1やSUMO2/3は、減数第一分裂終期でセントロメア局在が増加し、第二分裂中期にかけて減少していくのが分かる。スケールバーは5マイクロメートル(μm、1μmは1,000分の1mm)
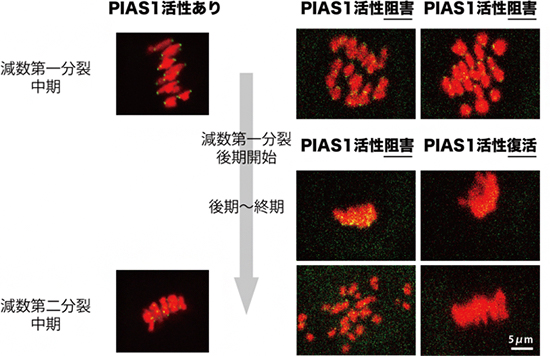
図3 SUMO化の機能阻害により失われるセントロメア接着
左: 正常な減数分裂では、第一分裂中期に相同染色体ペアが分裂面に並び、第二分裂中期では姉妹染色分体ペアが並ぶ。
中: PIAS1の酵素活性を阻害すると、第一分裂中期の染色体の配置は正常だが、第二分裂中期で姉妹染色分体が分離した。
右: 一方、第一分裂の後期開始後にPIAS1の酵素活性を復活させると、減数第一分裂を通してセントロメア接着が維持され、姉妹染色分体の異常な分離は見られなくなった。
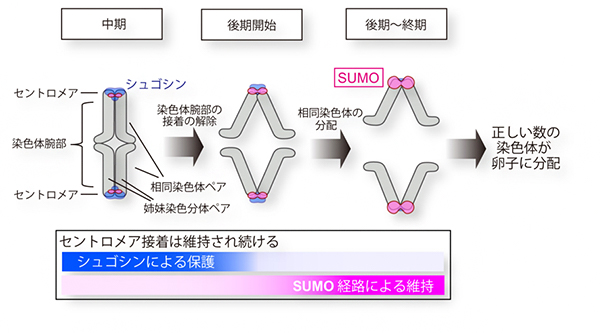
図4 減数第一分裂でのセントロメア接着を保護するシュゴシンとSUMO経路
減数第一分裂の後期に染色体腕部での姉妹染色分体の接着が解除されても、シュゴシンがセントロメア接着を保護している。シュゴシンがなくなった後はSUMO経路がセントロメア接着を維持する。