
2023-10-13 京都大学,基礎生物学研究所,東京理科大学
概要
植物は、減数分裂で生じた単相(n)の細胞から、多細胞体である「配偶体」をつくり、その中から生殖細胞を分化させるという、ヒトなどの動物とは異なる有性生殖の様式をもちます。配偶体は、花の咲く植物(被子植物)では花粉・胚のうという小さな組織ですが、コケ植物では葉状体・茎葉体という生活環の主役です。配偶体の中から生殖細胞が生じるメカニズムは、未だ多くの部分が不明のままになっています。
京都大学大学院生命科学研究科 齊藤美咲 修士課程学生(研究当時)、樅木亮介 修士課程学生(研究当時)、吉竹良洋 助教、宮川拓也 准教授、中野雄司 教授、荒木崇 教授、河内孝之 教授、山岡尚平 准教授らのグループは、海老根一生 基礎生物学研究所 助教、西浜竜一 東京理科大学 教授(元京都大学大学院生命科学研究科准教授)、光田展隆 産業技術総合研究所 生物プロセス研究部門 副研究部門長らと共同で、ほぼ全ての植物がもつ2つの遺伝子BONOBOとLRL/DROPが、生殖細胞の分化のために協調して働くことを明らかにしました。これらの遺伝子から生じるタンパク質は、組み合わさることで1つの複合体(ヘテロ二量体)を形成して他の遺伝子の発現を調節していました。LRL/DROPはシャジクモ藻類も持っていますが、BONOBOは陸上植物だけが持つ遺伝子です。陸上植物はおよそ5億年前にシャジクモ藻類の一種から誕生して進化してきましたが、このヘテロ二量体は、その頃に生み出され、進化の中で植物の生殖細胞をつくるための「鍵」として働いてきたと考えられます。
本成果は、2023年9月29日に国際学術誌「Current Biology」オンライン版に掲載されました。

1.背景
地球上の真核生物の多くは、雌雄それぞれの個体の中で減数分裂を行い、ゲノム1組だけをもつ単相(n)の細胞を生み出し、それらが卵と精子(配偶子)になり受精することで、両親由来のゲノム2組をもつ複相(2n)の細胞を生み出す、というライフ・サイクル(生活環)をもちます。ヒトを含む動物の多くは、単相の細胞全てがそのまま配偶子へと変化し、受精により複相の個体を生み出します。一方、陸上植物(被子植物・裸子植物・シダ植物・コケ植物など)では、単相の細胞は分裂を繰り返して「配偶体」とよばれる多細胞体となり、その中の一部の細胞のみが配偶子へと分化するという特徴があります(図1)。
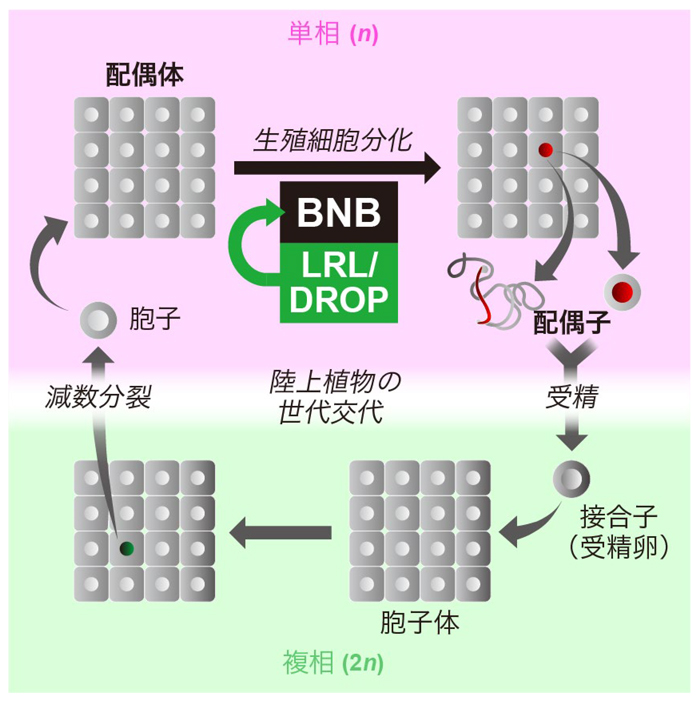
図1 陸上植物の世代交代と生殖細胞の分化
陸上植物は、単相(n)と複相(2n)のどちらの世代でも多細胞体を形成する。単相の多細胞体は「配偶体」と呼ばれ、その中の細胞から、のちに配偶子(卵・精子(精細胞))となる細胞(赤い丸をもつ細胞)が分化する。BNBとLRL/DROPのヘテロ二量体は、この生殖細胞分化のプロセスを制御する。
しかしながら、配偶体の様子は、被子植物とコケ植物で大きく異なります。被子植物の配偶体は細胞の数が非常に少なく、特に雄性配偶体である「花粉」は3細胞のみからなり、そのうちの2細胞が配偶子(精細胞)です。一方、コケ植物の配偶体は、私たちがふだん目にする植物個体(茎葉体・葉状体)であり、その中に造卵器・造精器を形成して配偶子をつくります。
これまでにわれわれの研究グループは、モデル植物※1であるゼニゴケの解析から、配偶体の中から配偶子を分化させる転写因子※2をつくる遺伝子BONOBO (BNB)を発見し、それが造卵器・造精器の元となる細胞(配偶子器始原細胞)を分化させる遺伝子であること、また被子植物シロイヌナズナがもつ相同遺伝子は、花粉の精細胞の元となる雄原細胞を分化させるために必要であることを明らかにしていました。BNBタンパク質は、塩基性ヘリックス・ループ・ヘリックス(bHLH)※3と呼ばれる構造をもち、二量体※4を形成することが予想されましたが、そのパートナーの分子は不明でした。
2.研究手法・成果
本研究では、シロイヌナズナにあるほぼ全ての転写因子(1,736個)の中でBNBタンパク質と結合するものを探索し、3個の転写因子(LRL1/DROP1、LRL2/DROP2、LRL3/DROP3)のみがBNBに結合し、ヘテロ二量体を形成することを見出しました。これらの遺伝子はbHLHサブファミリーXIという遺伝子ファミリー※5に属しており、ほぼ全ての陸上植物が相同遺伝子を持っていました。特に、一連の研究の出発点となったゼニゴケは、1つだけ相同遺伝子(MpLRL)を持っていました。またこれまでに、被子植物のLRL/DROPは根毛や根の構造の形成に関わり、MpLRLをはじめとするコケ植物の相同遺伝子は仮根の形成に関わることが知られていました。本研究において、シロイヌナズナの2つのLRL/DROP遺伝子をゲノム編集※6により同時に欠失させたところ、BNB遺伝子を失った場合と同様に、花粉において雄原細胞が分化しなくなりました。また、ゼニゴケでは、MpLRLの欠失により、配偶子器始原細胞の分化と、傘状の生殖器(雌器托・雄器托)の発生が見られなくなりました。このとき、ゼニゴケ、シロイヌナズナともに、配偶体においてBNB自体も発現しなくなったことから、LRL/DROPはBNBとのヘテロ二量体のパートナーであるだけでなく、BNBの安定な発現にも必要であることが分かりました(図2A-C)。これらのことから、LRL/DROPは、植物の生活環の中で、様々な細胞の分化に関わっており、有性生殖においてはBNBとヘテロ二量体を形成することで生殖細胞の分化を制御していると考えられます。bHLHサブファミリーXIの遺伝子は陸上植物だけでなくシャジクモ藻類も持っていますが、BNBが属するbHLHサブファミリーVIIIaファミリーの遺伝子は陸上植物しか持っていないことから(図2D)、陸上植物は、5億年前にシャジクモ藻類から分かれて誕生した頃にBNB-LRL/DROPヘテロ二量体を生み出し、配偶体から生殖細胞を分化させるために用いるようになったと考えられます。
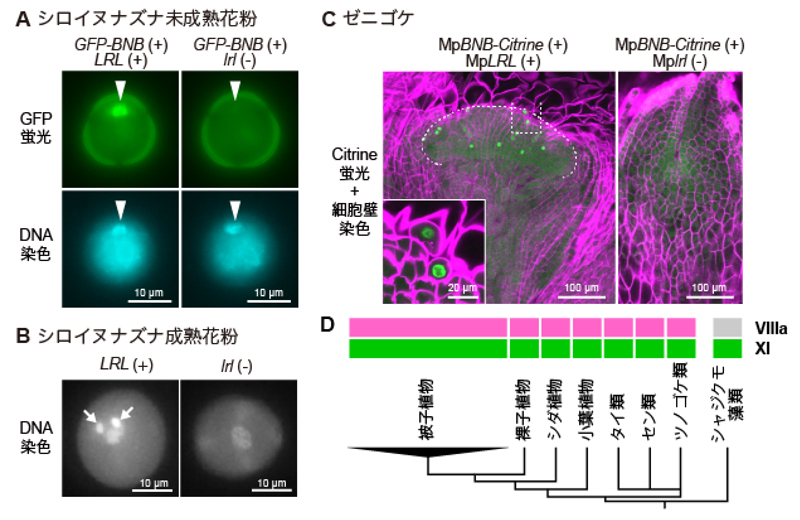 図2 BNB-LRL/DROPヘテロ二量体と陸上植物の進化
図2 BNB-LRL/DROPヘテロ二量体と陸上植物の進化
(A) シロイヌナズナの未成熟の花粉の中にできる雄原細胞(矢じり)では、LRL/DROPがあれば、蛍光タンパク質GFPで標識したBNBが発現するが(左:LRL (+))、LRL/DROPを欠失するとGFP-BNBも発現しない(右:lrl (-))。(B) 成熟した花粉では、LRL/DROPをもつ野生型では2個の精細胞(矢印)がみられるが(左:LRL (+))、LRL/DROPを欠失すると精細胞が失われる(右:lrl (-))。(C) ゼニゴケでは、MpLRLをもつ野生型株では、蛍光タンパク質Citrineで標識したBNBを発現する配偶子器始原細胞(緑色)が分化し、生殖器托の原基(点線で囲んだ部分)が発生するが(左:MpLRL (+))、MpLRLを欠失した変異株ではみられない(右:Mplrl (-))。(D) 陸上植物の系統樹とbHLH転写因子の分布。LRL/DROP の属するbHLHサブファミリーXI(緑)は陸上植物とシャジクモ藻類の両方がもつが、BNB の属するbHLHサブファミリーVIIIa(マゼンタ)は陸上植物に特有である。
3.波及効果、今後の予定
本研究により、植物の生活環とくに有性生殖の重要なプロセスの一つである生殖細胞の分化について、鍵となるメカニズムとその進化の一端が明らかになりました。今後は、BNB-LRL/DROPがどのようなメカニズムにより発現して生殖細胞を生み出すのかを詳しく調べることで、陸上植物の配偶子形成の全容とその進化の過程が解明されると期待されます。こうした植物の有性生殖のプロセスの解明は、農業・育種における新しい技術の開発やバイオテクノロジーの発展につながると期待されます。
4.研究プロジェクトについて
本研究は科学研究費補助金 学術変革領域(B)「細胞運命操作による植物生殖システムのリモデリング」、新学術領域「植物新種誕生の原理」、新学術領域「植物発生ロジックの多元的開拓」、基盤研究(S)「陸上植物の性分化:遺伝的頑健性と可塑性のメカニズム」、武田科学振興財団ライフサイエンス研究助成、旭硝子財団研究奨励などの支援を受けて行われました。
<用語解説>
※1 モデル植物:植物における基本的な生命現象を分子レベルで理解するために、全ゲノム情報を明らかにし、遺伝子組換え(形質転換)を可能にした種のこと。被子植物ではシロイヌナズナやイネ、コケ植物ではヒメツリガネゴケやゼニゴケが知られている。ゼニゴケは、京都大学発のモデル植物であり、京都大学の研究グループをはじめとする国際共同研究により、2017年に全ゲノム情報がはじめて解明された。
※2 転写因子:DNAの塩基配列をRNAに写し取る「転写」を調節するタンパク質のこと。転写されたRNAのうちの一種メッセンジャーRNA (mRNA)は、その後アミノ酸配列に読み換えられ(翻訳)、タンパク質を生み出す。
※3 塩基性ヘリックス・ループ・ヘリックス (bHLH):タンパク質のアミノ酸配列のうち、リジンやアルギニンなどの塩基性アミノ酸を10個程度もち、続いてらせん状(ヘリックス)構造をつくる15個程度のアミノ酸が、6〜10個程度のアミノ酸(ループ)をはさんで2つ並んだ配列から生じる立体構造のこと。DNAに結合するとともに、二量体(※4)をつくる性質をもち、主に転写因子として働く。真核生物の間に広く存在する、進化的に保存されたタンパク質の立体構造。
※4 二量体:同じ種類の分子(ここではタンパク質)が2個結合することによりつくられる複合体のこと。同じ分子同士が結合したものをホモ二量体、異なる分子が結合したものをヘテロ二量体と呼ぶ。
※5 遺伝子ファミリー:生物の種の中と種の間において、相同性の高い配列をもつ遺伝子のグループのこと。類似の機能をもつことが多い。生物の進化の中で、祖先種で誕生した1つの遺伝子が、重複したのち少しずつ異なる配列をもつようになり、それが分かれた生物種の間で受け継がれることで、遺伝子ファミリーが形成されていくと考えられている。
※6 ゲノム編集:ゲノムDNAの一部の配列を欠失・挿入・置換させることにより、遺伝子の構造を変化させ、その機能を改変・欠失させる技術のこと。2012年に報告されたCRISPR/Cas9システムは、効率の高さと汎用性から主要なゲノム編集技術として広く用いられるようになった。
<研究者のコメント>
この研究は、「その構造からヘテロ二量体を形成して機能すると予測されるBNBタンパク質の『相方』は何か?」という疑問から出発しました。機能未知のものも含む多数の候補の中から見つかった「相方」は、意外にも、根や根毛、仮根形成に関する機能が既に知られていたLRL/DROPでした。つまりこの研究には、LRL/DROPが二量体形成の相手を変えることにより、根等の形成と生殖細胞分化という全く異なる現象を制御していることを明らかにしたという側面もあります。植物が5億年も前からこのような緻密な制御機構を有し、種を超えて現在まで維持していることは、大変興味深いことです。(齊藤美咲)
<論文タイトルと著者>
タイトル:A bHLH heterodimer regulates germ cell differentiation in land plant gametophytes(陸上植物の配偶体における生殖細胞分化を制御するヘテロ二量体)
著 者:齊藤美咲1, 樅木亮介1, 海老根一生2,3, 吉竹良洋1, 西浜竜一1,4, 宮川拓也1, 中野雄司1,
光田展隆5, 荒木崇1, 河内孝之1, 山岡尚平1
1京都大学 大学院生命科学研究科
2基礎生物学研究所 細胞動態研究部門
3総合研究大学院大学 先端学術院
4東京理科大学 創域理工学部 生命生物科学科
5産業技術総合研究所 生物プロセス研究部門
掲 載 誌:Current Biology DOI:10.1016/j.cub.2023.09.020
")