2024-03-06 東京大学
発表のポイント
◆種によってGnRH1/3のいずれかが発現するといわれていたニューロンで、ピラニアは両方の遺伝子が共発現することを発見した。さらに、ピラニアの特殊な発現パターンをメダカ・ゼブラフィッシュにおいて擬似的に再現した結果から進化のシナリオを推定した。
◆これにより、脳下垂体を制御して繁殖制御を司るGnRHパラログが種ごとに異なる理由を初めて実験的に説明することができた。
◆理論的には比較的早期に冗長性が失われるとされる遺伝子重複後のパラログにおいて冗長性が保たれる例外を示し、遺伝子進化一般のセオリーと実態を繋げることが期待される。

ピラニア・ナッテリー
概要
東京大学 大気海洋研究所 海洋生命科学部門の神田真司准教授による研究グループは、GnRH神経系(注1)において発現し、排卵制御の鍵となるGnRHパラログ(注2)の発現に種間差が存在することについて、共通祖先が比較的最近まで保持していた冗長な発現が原因である可能性が高いことを明らかにしました(図1)。
機能的に相同な細胞においても、種によって発現するパラログが異なることが稀に見られますが、その理由はほとんどのケースで明らかにされておりません。本研究では、繁殖に必須と考えられているGnRH神経系をモデルとして、ピラニアにおけるGnRHパラログの発現を解析しました。すると、同じ機能のGnRHパラログ(gnrh1、gnrh3)が視床下部ニューロンで共発現し、両者が冗長的に脳下垂体機能を調節していることがわかりました。さらに、遺伝子ノックアウトや、他魚種でのエンハンサー(注3)活性解析でさらなる検証を行い、共通祖先がもっていたこの共発現が、現生種で脳下垂体制御に用いられているGnRHパラログのサブタイプが種間で一定しない原因となっていることを証明しました。この研究成果は今後遺伝子進化一般の理解に役立つことが期待されます。
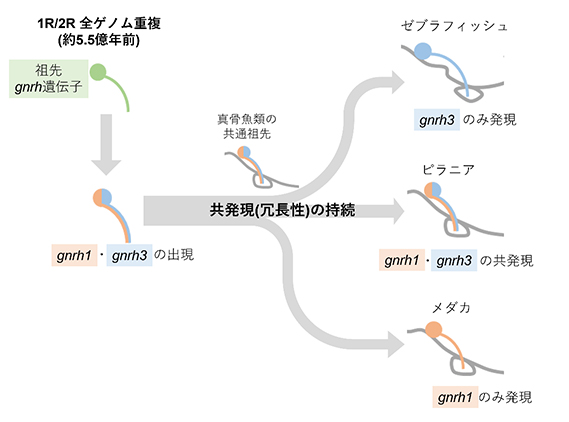
図1:本研究結果から提案された作業仮説
gnrh1とgnrh3の遺伝子重複後、ピラニアで現在も残されているように、両者の視床下部ニューロンにおける共発現は非常に長い期間で保たれていた。現生の様々な種でgnrh1とgnrh3が不規則的に脳下垂体制御に使われている理由は、それらの共通祖先が比較的最近までピラニアのような共発現を保っていたことによると示唆される。
発表内容
脊椎動物において視床下部に存在するGnRH神経系が脳下垂体の黄体形成ホルモン(LH)を放出させることによって排卵を引き起こすことが知られています。しかし、その視床下部GnRH神経系が発現するGnRH遺伝子は、GnRH1であったり、GnRH3であったり、種によって異なるパラログが用いられていることが長い間謎とされていました。そこで本研究では、カラシン目という系統群をモデルにした解析を行いました。カラシン目は、多くの種がgnrh3遺伝子のみを持つのに対し、ピラニアを含む一部の種はgnrh1、gnrh3の2つを保持する特徴を持つため、進化の途上でgnrh1を失うことが許容された理由を解析することができます。ピラニアの視床下部GnRH神経系を解析してみると、gnrh1とgnrh3が同じニューロンで共発現しており、いずれの免疫陽性線維も脳下垂体に同様に投射していたことから、GnRH1とGnRH3のパラログの両方が脳下垂体制御において冗長的に働いていることが強く示唆されました。gnrh1とgnrh3遺伝子は、5億年程度前におこった1R/2R全ゲノム重複(注4)で生じたと考えられており、比較的速やかに冗長性が解消される進化の過程でこれほどの長い期間冗長性が保たれていたことは非常に驚く結果でした。
本研究ではさらにピラニアのgnrh1、gnrh3エンハンサーの制御下で蛍光タンパク質を発現させるコンストラクトを導入したトランスジェニックゼブラフィッシュ、メダカを作製しました。これによって、祖先的形質であると推定されたピラニアの遺伝子がもしゼブラフィッシュ・メダカにおいて存在した場合、どこに発現するかを推定することができます。この実験の結果、GnRH3しかもたないゼブラフィッシュ、GnRH1のみが脳下垂体制御に関与するメダカのいずれにおいても、ピラニアのgnrh1、gnrh3エンハンサーは脳下垂体に投射するGnRHニューロンに蛍光タンパク質を発現させる活性を示しました(図2)。この結果は、GnRHニューロン自体は片方のパラログを失った後も、古来に祖先が失ったパラログも含め、両方のパラログを発現させる転写因子セットを保持していることを示唆します。したがって、これらのケースでは、転写因子発現がどちらかの遺伝子だけを発現させるように変化したのではなく、いずれかの遺伝子のエンハンサーあるいは遺伝子自体が失われたことによって脳下垂体制御に使うパラログが決まったことになります。
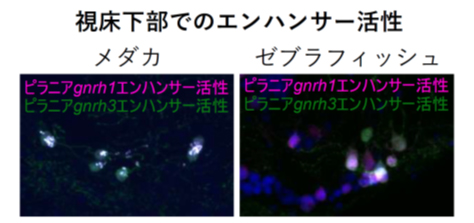
図2:ピラニアのgnrh1、gnrh3エンハンサーは、
メダカ(左)・ゼブラフィッシュ(右)の同一ニューロンに発現誘導活性を示す
メダカやゼブラフィッシュはそれぞれgnrh1あるいはgnrh3しか視床下部ニューロンで発現しないが、ピラニアのgnrh1、gnrh3エンハンサーは同じニューロンにおいて活性化された。すなわち、これらの種では、エンハンサー活性、あるいは遺伝子そのものが失われたことが、共発現をなくした理由であり、転写因子自体は両方のエンハンサーを活性化する能力を保持している。
これまで脳下垂体制御に用いられるGnRHパラログが、種によって異なることが広く認識されていましたが、本研究では現在に残された稀な共発現モデルを発見することにより、その原因を説明するひとつの合理的なモデルを提唱しました(図3)。種によって機能的に相同な細胞で発現する遺伝子サブタイプが異なる例は他にも存在するため、本研究で提示した作業仮説は、その他の遺伝子における進化を説明できるモデルとなり得るかもしれません。
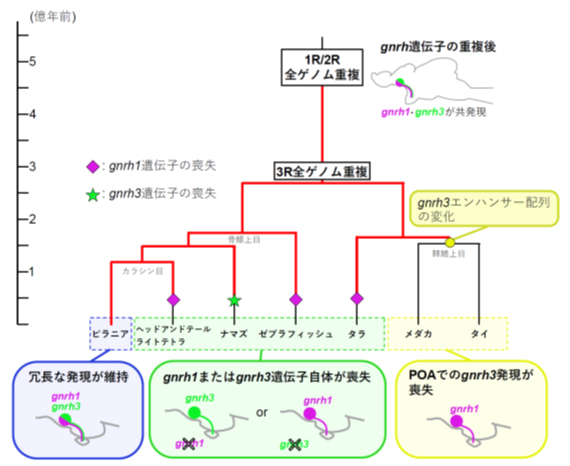
図3:真骨魚類の種によって、視床下部GnRH神経系に存在するgnrhサブタイプが異なるのは、
長く続いた共発現によると考えられる
それぞれのグループの遺伝子そのものの喪失あるいはエンハンサーの喪失により、共通祖先で共発現していたもののひとつが発現しなくなっている。ピラニアの含まれるグループは、現在もこの共発現を保っている非常に稀なケースである。
発表者・研究者等情報
東京大学 大気海洋研究所
神田 真司 准教授
藤森 千加 研究当時:特任研究員
現:北海道大学理学研究院 助教
杉本 航平 研究当時:修士課程
論文情報
雑誌名:iScience
題 名:Long-lasting redundant gnrh1/3 expression in GnRH neurons enabled apparent switching of paralog usage during evolution
著者名:Chika Fujimori, Kohei Sugimoto, Mio Ishida, Christopher Yang, Daichi Kayo, Soma Tomihara, Kaori Sano, Yasuhisa Akazome, Yoshitaka Oka, Shinji Kanda*
DOI:10.1016/j.isci.2024.109304
URL:https://www.sciencedirect.com/science/article/pii/S258900422400525X![]()
研究助成
本研究は、科研費「挑戦的萌芽研究(課題番号:24657050)」、「挑戦的研究(萌芽)(課題番号:18K19323)」、「新学術領域研究(公募研究)(課題番号:18H04881)」、「基盤研究(B)(課題番号:23H02306)」、三菱財団自然科学研究助成、住友財団 基礎科学研究助成、三島海雲記念財団 学術研究奨励金の支援により実施されました。
用語解説
- (注1)GnRH神経系
- 生殖腺刺激ホルモン放出ホルモンとよばれるGnRHを産生し、放出する神経系。視床下部に存在するGnRH神経系は、脳下垂体の生殖腺刺激ホルモンの放出を制御することにより、卵胞の発育や排卵、あるいは精子形成などの生殖腺の機能を調節する。
- (注2)パラログ
- 新しい遺伝子は多くの場合遺伝子重複(コピー)によって生じる。パラログとは、同じ遺伝子から生じた兄弟の遺伝子のこと。
- (注3)エンハンサー
- 遺伝子の発現制御に関するDNA配列。細胞は、保持する転写因子によって特定のエンハンサーを選択的に活性化することで遺伝子発現を制御する。このため、遺伝子発現の制御にはエンハンサーと転写因子の組み合わせが重要なはたらきをする。
- (注4)1R/2R全ゲノム重複
- 全ゲノム重複は、全ゲノムが染色体レベルでそのまま倍になる現象。多くの遺伝子が同時にコピーされるため、進化の駆動力となると考えられている。1R/2R全ゲノム重複は、脊椎動物の誕生時、および無顎類が分岐したあとにおこったと考えられる、二回の大規模な全ゲノム重複のことで、哺乳類を含む四肢動物・硬骨魚類・軟骨魚類に共通する遺伝子セットを作り出した。
問合せ先
東京大学大気海洋研究所 海洋生命科学部門
准教授 神田 真司(かんだ しんじ)

