
2024-03-15 東京大学
発表のポイント
◆結核菌(Mycobacterium tuberculosis)が保有する、自身の転移RNA(tRNA)を不活性化し、増殖抑制作用をもつMenT3トキシン(毒素)タンパク質の基質特異性を解明しました。
◆MenT3はセリンを受容するtRNAを特異的に認識し、その3’末端に連続したシチジン(cytidine)配列を付加することが判明しました。
◆結核菌のトキシンタンパク質の活性を阻害し、結核菌に対する抗生物質をより効果的に作用させる新しいタイプの薬剤開発が期待されます。

結核菌のMenT3トキシンタンパク質によるtRNAの不活性化と特異性の分子機構
発表概要
東京大学大学院新領域創成科学研究科メディカル情報生命専攻の劉俊大学院生(博士課程)と富田耕造教授らは、ヒトにおいて肺結核を引き起こす結核菌(Mycobacterium tuberculosis)が保有するMenT3トキシンタンパク質の基質特異性の分子機構を解明しました。
細菌は栄養飢餓状態や抗生物質暴露などの環境ストレス下での生存を可能にするトキシン(毒素)-アンチトキシン(抗毒素)システム(注1)を保持しており、ストレス存在下では、自身のトキシンによって増殖を抑制し、環境ストレスから逃れます。
MenT3トキシンタンパク質は、特定の転移RNA(transfer RNA;tRNA、注2)の3’末端に特定のヌクレオチド(注3)を付加することによりtRNAを不活性化し、タンパク質合成を阻害し、自身の増殖を抑制する機能があります。
本研究成果は結核菌の増殖制御に関与するMenT3トキシンタンパク質の活性を阻害し、結核菌に対する抗生物質をより効果的に作用させる新しいタイプの薬剤開発に寄与することが期待されます。
本研究成果は、3月15日に『Nucleic Acids Research』誌にオンライン掲載されました。
発表内容
細菌が有するトキシン(Toxin)-アンチトキシン(antitoxin)対(TAモジュール)(注1)は、細菌が多様な環境ストレス条件下で生存していくために獲得してきたシステムであり、一方で、細菌の病原性や薬剤耐性などに関与しています。
結核菌(Mycobacterium tuberculosis)はヒトにおいて肺結核を引き起こす病原性細菌であり、空気感染により広がり、感染後、慢性的な呼吸器症状を引き起こします。最近、結核菌において、新たなTAモジュールが見いだされました。このTAモジュールのトキシンタンパク質(MenT3)は転移RNA(transfer RNA;tRNA)の3’末端にヌクレオチドを付加し、細菌内のタンパク質合成を阻害し、結核菌の増殖を抑制します。これまで、MenT3がtRNAの3’末端に付加するヌクレオチドの特異性やMenT3が標的とする細菌内のtRNA分子種、またその特異性の分子機構は不明でした。
本研究では、まず、MenT3がCTP(cytidine triphosphate:シチジン(cytidine、注4)・三リン酸)を優先的に基質として用いて、tRNAの3’末端に連続したシチジン配列を付加することが明らかになりました。また、MenT3とCTPの複合体X線結晶構造解析から、MenT3のヌクレオチド結合ポケットの形状がCTPに適したものであり(図1左)、他のヌクレオチドは塩基部分がそのポケットと相互作用できないことも明らかになりました(図1右)。
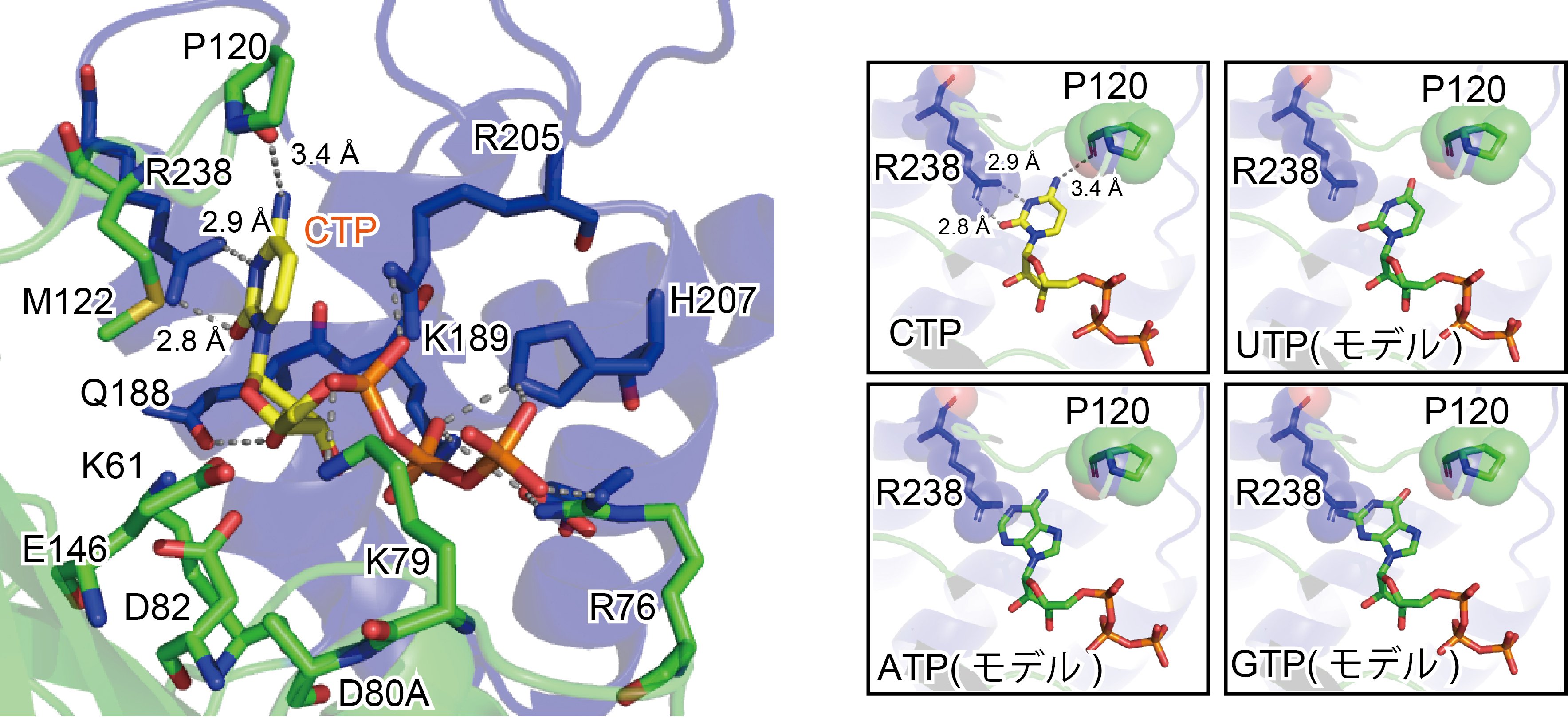
図1:MenT3とCTPの複合体構造
(左)MenT3のヌクレオチド結合ポケットはCTPに適した大きさと形をしており、また、CTPの塩基部分はポケット内のアミノ酸(Pro120の主鎖およびArg238の側鎖)によって認識されている。
(右)UTP、ATP、GTPとの結合モデル。CTPとの結合と比較。
次に、MenT3の作用により、細胞内ではセリンを受容するtRNA(tRNASer)種のみが著しく減少することが明らかとなり、また、tRNASerに特徴的な長いバリアブル・ループ(variable loop, V-loop)がMenT3によるtRNASerへのシチジン付加に必要であることも明らかになりました(図2)。
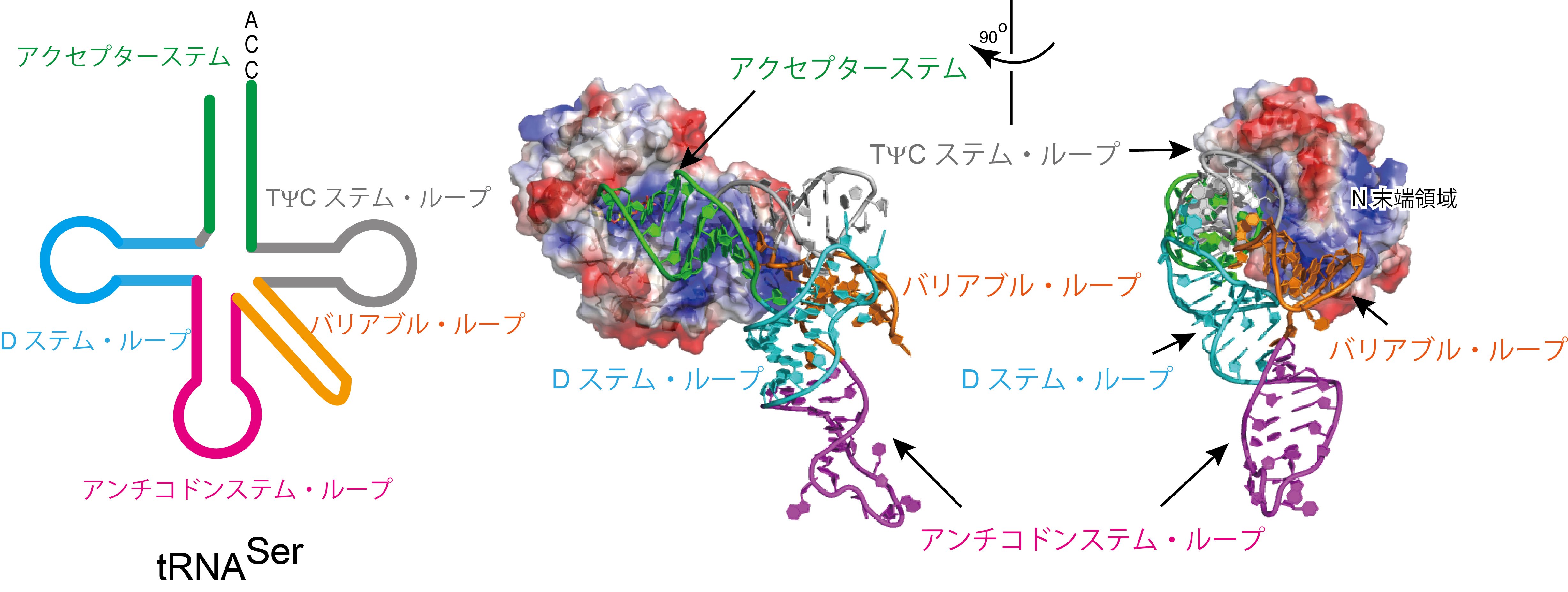
図2:MenT3は長いバリアブル・ループをもつtRNASerを不活性化
(左)tRNA分子の2次構造。MenT3によるtRNASerの3’末端へのシチジン付加にはtRNASerの有する特徴的な長いバリアブル・ループ(オレンジ色部分)を必要とする。
(右)MenT3(表面電荷表示)とtRNASerの複合体モデル。プラス表面電荷(青)およびマイナス電荷(赤)。バリアブル・ループがMenT3のN末端領域の正電荷部位と相互作用。
さらに、MenT3のアミノ末端(N末端)にある正電荷領域がtRNASerのv-loopを認識していることも明らかになりました。以上より、MenT3は長いv-loopをもったtRNASerを選択的に認識し、その3’末端にシチジンを付加し、tRNASerを不活性化するユニークなトキシンタンパク質であることが明らかになりました。
国内で検出される多くの結核菌は、従来使用されていた抗生物質であるストレプトマイシンに対してすでに耐性を持っています。そのため、現在では主にイソニアジドとリファンピシンを中心とした治療が行われています。しかし、これらの薬剤に耐性のある多剤耐性結核菌も見つかっており、新しい治療法の開発が求められています。結核菌が抗生物質暴露などのストレス条件下で生存していくために獲得してきた増殖制御機構を担うトキシンタンパク質の機能を阻害することによって、結核菌に対する抗生物質がより効果的に作用する可能性があります。本研究成果は結核菌に対する新しいタイプの薬剤開発に寄与することが期待されます。
発表者
東京大学大学院新領域創成科学研究科 メディカル情報生命専攻
劉 俊(博士課程)
富田 耕造(教授)
論文情報
雑誌名:Nucleic Acids Research
題名: Substrate specificity of Mycobacterium tuberculosis tRNA terminal nucleotidyltransferase toxin MenT3
著者名:Jun Liu, Yuka Yashiro, Yuriko Sakaguchi, Tsutomu Suzuki and Kozo Tomita*
DOI: 10.1093/nar/gkae177
URL: https://doi.org/10.1093/nar/gkae177
研究助成
本研究は、日本学術振興会科学研究費助成事業 基盤研究A(23H00368、18H03980)、武田科学振興財団 ハイリスク新興感染症研究助成などの支援を受けて行われました。
用語解説
(注1)トキシン(毒素)-アンチトキシン(抗毒素)システム
毒素(トキシン、Toxin)と抗毒素(アンチトキシン、Antitoxin)の遺伝子対から構成されています。細菌のTAモジュールのトキシンタンパク質は、DNA複製、転写、タンパク質合成、細胞壁合成などの重要な細胞内プロセスを妨げ、それによって細菌の成長を抑制、制御しています。通常の生理条件下では、アンチトキシン(RNAまたはタンパク質)がトキシンの発現やその活性を抑制することで、トキシンの毒性を中和しています。環境ストレス条件下では、アンチトキシンは分解または発現が抑制され、その結果、アンチトキシンによるトキシンの発現抑制や活性の抑制が解除され、細菌は自身の増殖を抑制、停止します。
(注2)tRNA
転移RNA(transfer RNA)。通常76-90ヌクレオチドからなるRNA分子であり、タンパク質をコードするメッセンジャーRNA(mRNA)とそのアミノ酸配列とを結びつけるアダプター分子として働きます。tRNAの3’末端にはCCA配列があり、その3’末端にアミノアシル合成酵素によってアミノ酸が付加されます。
(注3)ヌクレオチド
DNAやRNAを構成する単位であり、塩基部分とリボースそしてリン酸基からなります。ヌクレオチドが連なったものはポリヌクレオチドと呼びます。RNAは塩基部分が4種類 -アデニン(A)、グアニン(G)、ウラシル(U)、シトシン(C)-のヌクレオチドを構成成分として用い、またそれぞれのヌクレオチド・三リン酸化合物はATP、GTP、CTP、UTPと呼びます。
(注4)シチジン
RNAは主にアデノシン、グアノシン、ウリジン、シチジンの4種類のヌクレオチドを構成成分として用いています。
お問い合わせ
新領域創成科学研究科 広報室

")