
2024-07-19 基礎生物学研究所,奈良先端科学技術大学院大学,北海道大学,関西学院大学,理化学研究所,愛知教育大学
マメ科植物は根粒菌と共生することで、窒素栄養が乏しい土壌環境でも生育できます。これは根粒菌が大気窒素をアンモニアに変換する能力(窒素固定能)を持ち、マメ科植物は根粒菌の働きを通して大気中の窒素を利用できるからです。このような有益な細菌をマメ科植物は根に形成するコブ状の根粒に共生させています。しかし、無秩序に多くの根粒が形成されると土壌から吸収した水や他の栄養を葉や茎に輸送するといった根の本来の機能を損なう恐れがあります。そのため、宿主となるマメ科植物は根粒菌の感染によって根粒の分布が過密に形成されないように調節する必要がありますが、その機構は分かっていませんでした。
今回、マメ科のモデル植物であるミヤコグサの研究から、根における根粒菌への応答には一定のリズムを刻む周期的な遺伝子の働きが伴っており、その周期性が根粒菌の感染を許す根の領域の広さを規定することで、根粒の分布を調節していることを発見しました。さらに、この遺伝子発現のリズムの維持には植物ホルモンであるサイトカイニンが必要なことを明らかにしました。この研究成果は、基礎生物学研究所 共生システム研究部門 征矢野敬准教授、川口正代司教授、大熊直生特任助教(現 理化学研究所)、奈良先端科学技術大学院大学 先端科学技術研究科 中島敬ニ教授、郷達明助教、北海道大学大学院 理学研究院 生物科学部門 綿引雅昭准教授、関西学院大学 生命環境学部 武田直也教授、赤松明助教(現 理化学研究所)、理化学研究所 環境資源科学研究センター 質量分析・顕微鏡解析ユニット 榊原均客員主管研究員、小嶋美紀子専門技術員、竹林裕美子テクニカルスタッフI、愛知教育大学 教育学部 菅沼教生名誉教授らの共同研究により得られたものであり、米国の科学誌 Scienceに2024年7月19日に掲載されます。

【研究の背景】
根粒菌の感染は主にマメ科植物の根の先端部に近い狭い領域(高感受性領域)で起こります(図1A)。この領域よりもさらに根端側には根を構成する新しい細胞を生み出す根端分裂組織があるため、高感受性領域は根の成長に伴って継続的に形成されます。根粒菌の感染が常に高感受性領域で同程度の頻度で起こるのなら、根のどの部分でも感染部位は均等に分布するはずです。しかし、感染部位の分布をよく見ると密に形成された場所とそうでない場所が交互に現れる特徴があります(図1C, D)。また、根粒菌に応答して速やかに発現上昇する宿主遺伝子(NSP1遺伝子)も発現する場所としない場所が交互に現れます(図1B)。このことは高感受性領域での根粒菌に対する応答が断続的に繰り返し起こることを示唆します。しかし、根粒菌に対する根の応答動態を時系列に沿って詳しく調べた報告はありませんでした。
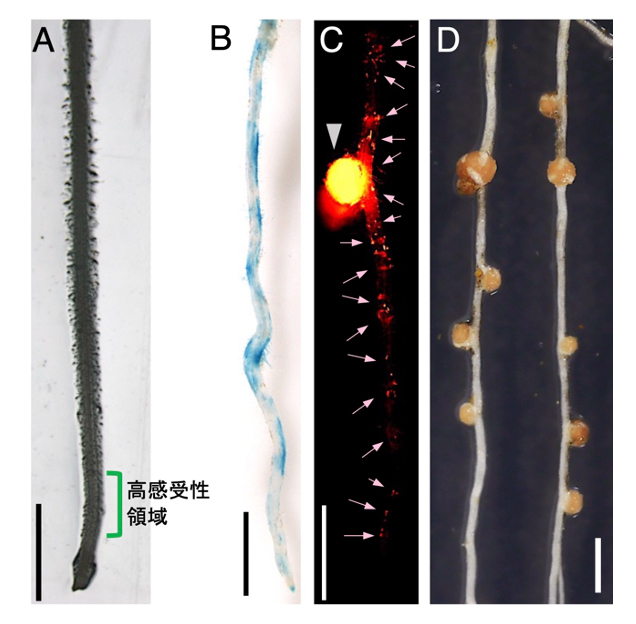
図1. ミヤコグサ根粒共生の初期に応答する宿主遺伝子(NSP1)の発現パターンと根粒菌感染部位の分布
(A)根端を含むミヤコグサの根。根粒菌に対して高感受性を示す領域を緑のラインで示した。(B)GUS染色で検出した根粒菌接種後のNSP1の発現。(C)蛍光標識した根粒菌を接種したミヤコグサの根。根粒を矢頭で、感染部位を矢印で示した。(D)根粒の分布。スケールバー: 2mm
【研究の成果】
研究チームは、根粒菌感染に必須な遺伝子として知られ、根粒菌に対して速やかに発現が上昇することが知られているNSP1遺伝子に注目し、高感度でかつ高時間分解能で遺伝子発現を検出することが可能な発光ライブイメージングを用いてNSP1遺伝子の時空間的な発現パターンを観察しました(図2A)。NSP1は、根粒菌を接種した後に、高感受性領域において約6時間の周期で発現が増減していること(図2B)、および、根の生長に伴い新たなNSP1発現領域が前の周期に形成された発現領域の先端部側に断続的に繰り返し形成されることが分かりました(図2A)。また、NSP1の周期的発現が強く見られた領域で根粒のほとんどが形成されたことから、この発現が根粒形成プロセスと密接な関係にあることが示唆されました。さらに、NSP1だけでなく、根粒共生に重要な多くの既往遺伝子の発現が周期性を示したことから、根粒菌に対する初期応答は周期的な遺伝子発現を伴う現象であることが初めて明らかになりました。
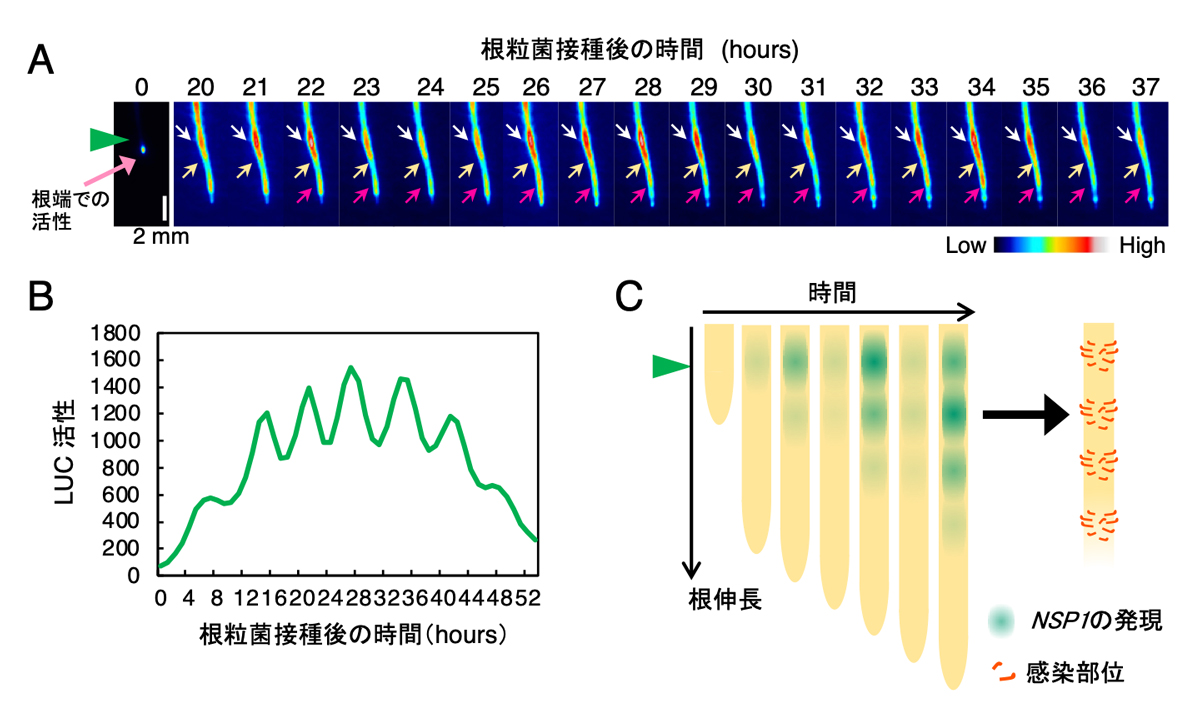 図2. NSP1遺伝子の周期的発現パターン
図2. NSP1遺伝子の周期的発現パターン
(A)根粒菌の感染に必須なNSP1遺伝子の時空間的発現パターン。白、黄、赤の矢印は断続形に形成されたNSP1の周期的発現量領域を示す。(B)矢頭で示した部位のNSP1の発現パターンをLUC活性によって経時的に測定した結果。(C)感染部位の関係を示した模式図。
周期的に発現する遺伝子には、植物ホルモンとして知られるサイトカイニンの生合成や代謝、情報伝達に関係するものが多く含まれていました。
そこで研究チームは根粒菌が感染する際のサイトカイニンに対する応答について詳細に調べました。サイトカイニンに対する応答のマーカーであるTCSnレポーターを用いて、サイトカイニン応答が活性化された場所を調べると、活性化された場所とそうでない場所が根の成長軸に沿って交互に観察されました(図3A)。TCSnの活性を発光ライブイメージングで捉えると、根粒菌接種後のサイトカイニンに対する応答も6時間周期で起こることが分かりました(図3B,C)。また、サイトカイニンの蓄積量の変化にも周期性が見られ、サイトカイニンの生合成や不活化を担う遺伝子の発現のタイミングもサイトカイニン応答と合致していました。
根粒共生においてサイトカイニンは重要な調節因子として働くことが知られています。例えば、サイトカイニンは根粒菌の感染には抑制的に作用することが分かっています。その一方で、サイトカイニンの蓄積には共生器官である根粒の形成を誘導するといった正の効果も知られています。サイトカイニン応答と遺伝子発現の周期性との関係を調べるために、サイトカイニン受容体であるLHK1の機能欠損変異体(lhk1)と活性化型遺伝子(gofLHK1)を導入した遺伝子組み換えミヤコグサを作成しました。サイトカイニン応答が活性化されたgofLHK1株では根粒菌接種時のNSP1の発現量が低下し、発現の周期性も乱されていました。この発現パターンは、サイトカイニンを直接与えた根に根粒菌を接種した場合とよく似ており、サイトカイニン応答が活性化すると共生関連遺伝子の発現が抑制されることが考えられました(図3D)。一方、サイトカイニンを受容できないlhk1変異体ではサイトカイニン応答が低下しており、NSP1の発現量は維持されていましたが(図3D)、発現周期の間隔と発現領域が拡大していました(図4A,B)。これらの結果より、サイトカイニン応答がNSP1の発現周期性の維持と発現領域の大きさの制御に必要なことが分かりました。
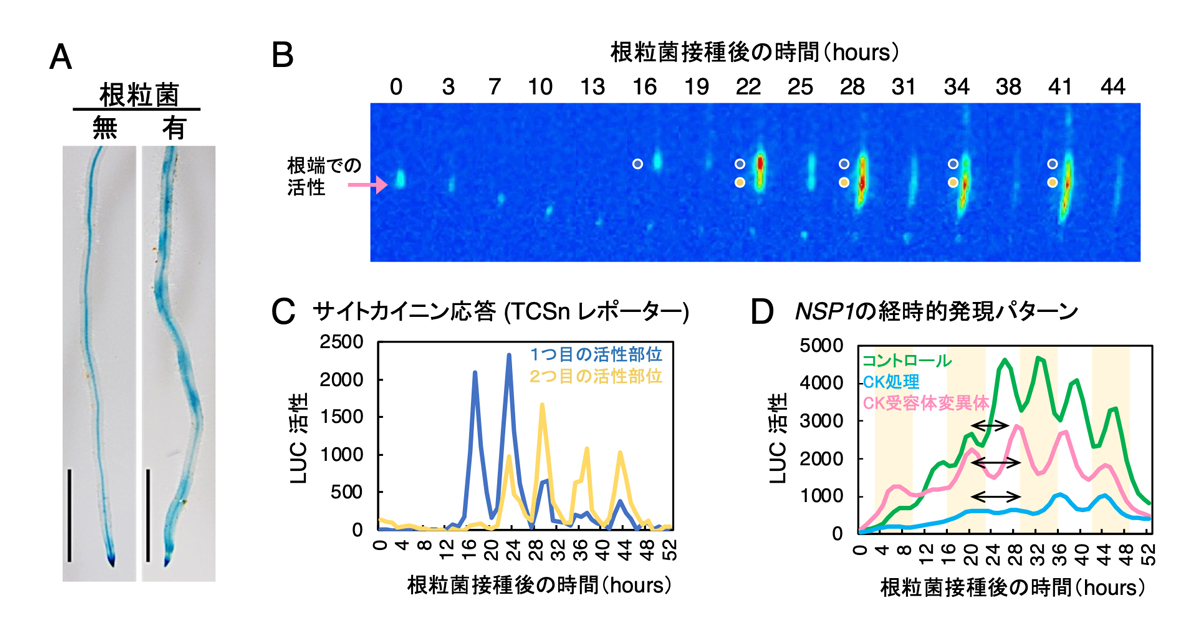 図3. サイトカイニン応答の周期的変化とサイトカイニンがNSP1の発現に及ぼす効果
図3. サイトカイニン応答の周期的変化とサイトカイニンがNSP1の発現に及ぼす効果
(A) GUS染色で検出したサイトカイニン応答。スケールバー: 2mm (B, C)LUC活性で可視化したサイトカイニン応答の経時的及び空間的発現パターンのライブイメージング(B)と、LUC活性測定結果(C)。(D)NSP1の発現に対するサイトカイニン (CK) 処理及びサイトカイニン受容体遺伝子の機能欠損の効果。
NSP1は根粒菌の感染に必須な遺伝子であり、その発現部位は感染部位が形成される領域と一致します。共生関連遺伝子の発現周期性と感染部位の分布の関連性を調べるために、感染部位が密に形成される領域を一つの単位としてその大きさを測定しました。lhk1機能欠損変異体では正常な株と比較して、その領域が拡大していました(図4C)。感染部位は増えるもののNSP1遺伝子の発現周期は変化しない別の変異体では、領域の大きさは正常な株と同程度であることも分かりました。一方、周期性が乱れるgofLHK1株では感染部位の数が減少しており、感染部位領域の大きさはやや拡大しながら全体としてばらつきが大きくなりました(図4C)。また、表皮での感染部位直下に形成される根粒の分布にも感染部位の分布と一致した変化が見られました。以上の結果から、サイトカイニンの正常な受容が共生関連遺伝子の発現周期性を維持しており、この経路が破綻するとこれらの遺伝子の発現領域の大きさが変化し、感染部位や根粒の分布に影響することが分かりました(図5)。
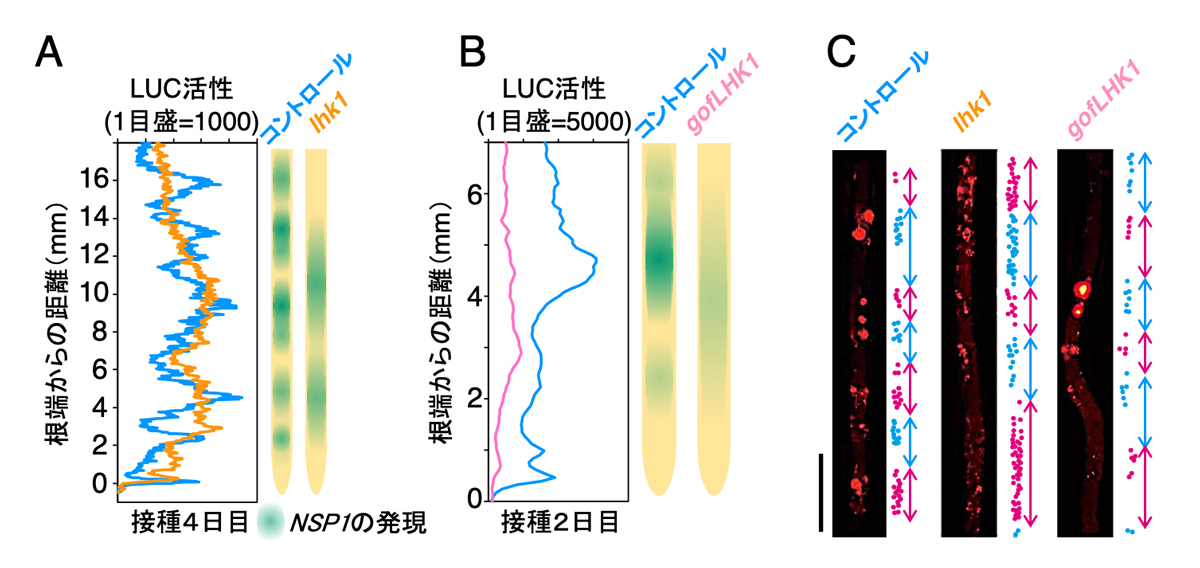 図4. サイトカイニン応答の周期的変化がNSP1の空間的発現パターンと感染部位の分布に及ぼす効果
図4. サイトカイニン応答の周期的変化がNSP1の空間的発現パターンと感染部位の分布に及ぼす効果
(A, B) 根の伸長軸に沿ったNSP1の発現パターン。コントロールとlhk1変異体(A)、 gofLHK1株(B)の比較。(C)感染部位の分布。ドットは感染部位の位置を示す。階層的クラスタリングでグループ分けした。スケールバー:2mm
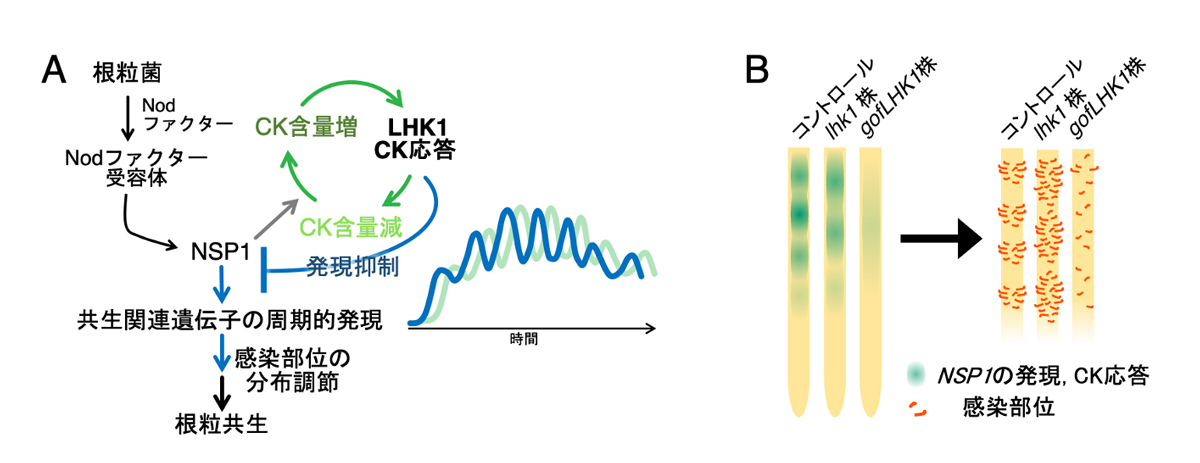 図5. サイトカイニン応答と共生関連遺伝子の発現周期性のモデル
図5. サイトカイニン応答と共生関連遺伝子の発現周期性のモデル
(A)根粒菌の感染によって共生経路が活性化され、周期的サイトカイニン応答が誘導される。サイトカイニンは共生関連遺伝子の発現に抑制的に作用するため、サイトカイニン応答が遺伝子発現の周期に影響する。(B) 周期性の変化が共生関連遺伝子の空間的発現領域の大きさに影響し、感染部位や根粒の分布パターンが変化する。
【今後の展望】
根粒共生は被子植物の中でも単系統群(窒素固定クレード)を構成するマメ目、バラ目、ウリ目、ブナ目で見られ、これらの共通祖先が根粒共生を獲得したと考えられています。その中でもマメ目マメ科はほとんどの種で根粒共生が見られます。マメ科以外の根粒共生ではサイトカイニンの作用はほとんど報告がなく、マメ科植物が独自にサイトカイニン経路を根粒共生の重要な制御要素として取り込んだ可能性があります。サイトカイニンに対する周期的応答は意外な発見であったため、この発見と同時に周期性を形成する分子機構や周期的応答が感染領域を形成する仕組みなど、今後追求すべき多くの課題が浮かび上がりました。また、植物ホルモンによる周期的な遺伝子発現制御はシロイヌナズナにおいてオーキシンが介在する系が知られているのみであり、この意味でも今回の発見は意義があると考えられます。今後、これらの課題を追求することで、根粒共生の制御機構の解明が進み、植物ホルモンを介した周期的反応による器官発達の空間制御機構についての理解も深まることが期待されます。
【発表雑誌】
雑誌名: Science 2024年7月19日号 掲載
掲載日: 米国東部時間7月18日14時(日本時間19日午前3時)にオンライン公開
論文タイトル:Periodic cytokinin responses in Lotus japonicus rhizobium infection and nodule development
著者:Takashi Soyano, Akira Akamatsu, Naoya Takeda, Masaaki K. Watahiki, Tatsuaki Goh, Nao Okuma, Norio Suganuma, Mikiko Kojima, Yumiko Takebayashi, Hitoshi Sakakibara, Keiji Nakajima, Masayoshi Kawaguchi
DOI:https://doi.org/10.1126/science.adk5589
【研究グループ】
自然科学研究機構 基礎生物学研究所 共生システム研究部門 征矢野 敬 准教授、川口 正代司 教授、大熊 直生 特任助教(現 理化学研究所 基礎科学特別研究員)
奈良先端科学技術大学院大学 先端科学技術研究科 中島 敬ニ 教授、郷 達明 助教
北海道大学大学院 理学研究院 生物科学部門 綿引 雅昭 准教授
関西学院大学 生命環境学部 生物科学科 武田 直也 教授、赤松 明 助教(現 理化学研究所 研究員)
理化学研究所 環境資源科学研究センター 質量分析・顕微鏡解析ユニット 榊原 均 客員主管研究員(兼 名古屋大学大学院 生命農学研究科 教授)、小嶋美紀子 専門技術員、竹林 裕美子 テクニカルスタッフI
愛知教育大学 教育学部 菅沼 教生 名誉教授
【研究サポート】
この成果は下記の研究サポートを受けて実施されました。
科学研究費助成事業 (19K05959, 22H02304, 22K06288)
科学研究費助成事業 新学術領域研究 「植物の周期と変調」 (20H05429, 22H04737, 19H05670, 19H05671, 19H05673)
公益財団法人 武田科学振興財団 生命科学研究助成
基礎生物学研究所 超階層生物学センター データ統合解析室、モデル生物研究支援室
理化学研究所 科技ハブ産連本部「合成生物学による次世代細胞農業-代謝モジュール細胞の確立とその人工組織化のためのプラットフォーム開発」
【本研究に関するお問い合わせ先】
基礎生物学研究所 共生システム研究部門
准教授 征矢野 敬(ソヤノ タカシ)
【報道担当】
基礎生物学研究所 広報室
奈良先端科学技術大学院大学 企画総務課 渉外企画係
北海道大学 社会共創部広報課広報・渉外担当
学校法人関西学院 広報部 企画広報課
理化学研究所 広報室 報道担当
愛知教育大学 学術研究支援課
")
