
2024-10-04 理化学研究所
理化学研究所(理研)開拓研究本部 伊藤ナノ医工学研究室(研究当時)の森島 信裕 客員研究員(研究当時、現 光量子工学研究センター 先端レーザー加工研究チーム 客員研究員)、伊藤 嘉浩 主任研究員(研究当時、現 光量子工学研究センター 先端レーザー加工研究チーム 客員主管研究員)の研究チームは、カルシウム依存性タンパク質分解酵素カルパイン[1]の一つ、カルパイン5が骨格筋[2]の形成過程で活性化し、筋タンパク質の合成や核の位置決めを制御することを発見しました。
本研究成果は、骨格筋形成の制御機構およびカルパインの生理機能の解明、さらにはカルパイン5が関わる病気(血管新生炎症性硝子体網膜症[3])の治療法開発に貢献すると期待されます。
今回、研究チームは、筋芽細胞(骨格筋前駆細胞)同士が融合して多核の骨格筋細胞になる過程[4]を調べ、融合の前にカルパイン5が一過的に活性化する現象を見つけました。カルパイン5は、細胞増殖や分化の制御にとって重要な転写調節タンパク質を切断します。遺伝子ノックダウン[5]法を用いてカルパイン5の量を減らすと、細胞融合後の多核細胞中で筋収縮タンパク質が十分作られず、筋力発生の源である筋原線維[6]構造が形成されませんでした。また、多数の核が細胞内に広がらずに凝集するという異常が生じました。これは、骨格筋形成にとってカルパイン5の働きが重要であることを示唆します。
本研究は、科学雑誌『Journal of Biological Chemistry』オンライン版(9月30日付)に掲載されました。

カルパイン5の働きと筋タンパク質合成、細胞中の核配置
背景
体を構成する組織や臓器は細胞分化によって作られます。細胞分化は前駆細胞の中のタンパク質をはじめとする構成成分や細胞内外の構造が変化することによってもたらされます。こうした変化は遺伝子発現(DNAが転写されてmRNAが作られ、さらにそれを基にしてタンパク質が作られる過程)を調節する多数の転写調節タンパク質(転写調節因子[7])の働きによってなされます。骨格筋形成もその一例で、筋分化特異的転写調節因子が中心的な働きをすることによって筋芽細胞(骨格筋前駆細胞)から筋肉細胞が作られることが知られています。転写調節因子はiPS細胞のような未分化細胞を使って人為的に組織、臓器を作り出す再生医療においても重要です。
筋芽細胞は分化誘導条件下で細胞分裂を停止し、多数の筋芽細胞同士が融合して筋細胞(多核の大きな細胞)が形成されます。増殖がストップすると、細胞融合が始まる前の段階にある筋芽細胞の中で小胞体[8]という細胞小器官からカルシウムが細胞質ゾル(細胞質の可溶性部分)へ流出し注1)、小胞体ストレス[9]が引き起こされます注2)。この現象が起こると補償作用として細胞は細胞外からカルシウムを取り込むため、細胞質ゾル中では小胞体と細胞外からのカルシウムによってカルシウム濃度が上昇します。細胞にはカルシウムが結合すると活性化するタンパク質分解酵素(カルパイン)が不活性な状態で常在していることから、研究チームは小胞体ストレスが起きる時期にカルパインが活性化し、筋分化過程の進行に何らかの役割を果たしているのではないかと考えました。ヒトやマウスのカルパインは15種類のメンバーから成ります。そのうちの一つ、カルパイン5は多くの器官、組織に存在しています。
注1)2015年2月17日プレスリリース「筋肉を動かすカルシウムは筋肉を作る指令役も担う」
注2)2005年5月23日プレスリリース「筋肉組織の構築に細胞ストレスが必要なことを発見(PDF 483.2KB)![]() 」
」
研究手法と成果
研究チームは筋分化研究において最もよく用いられている培養筋芽細胞の一つ、マウスC2C12細胞の筋分化過程を調べました。カルパインによるタンパク質切断は限定分解と呼ばれる様式で起こり、基質タンパク質中の特定配列を切断して基質を断片化します。研究チームは限定分解を解析するのに適しているウエスタンブロット法[10]を用いて、筋芽細胞中でカルパインの基質候補(STAT3、c-Myc)が切断されているかを探りました。カルパインによるタンパク質切断はこれまで主に病理的条件で見つかってきましたが、STAT3やc-Mycは増殖や分化の制御に関わる転写調節因子で、血小板など生細胞中でカルパイン1や2によって切断されることが知られていました。
筋分化過程は比較的ゆっくりしたスピードで進み、細胞増殖から細胞分化への切り替えに1、2日、さらに筋収縮を起こす筋細胞ができるまでには1週間程度の日数を要します。研究チームは、小胞体ストレスが起こる分化誘導開始翌日に1時間刻みで解析を行いました。その結果、C2C12細胞を分化誘導条件下において24時間から35時間の間に数回、それぞれ約1時間の間STAT3、c-Mycが切断されることを見いだしました(図1)。
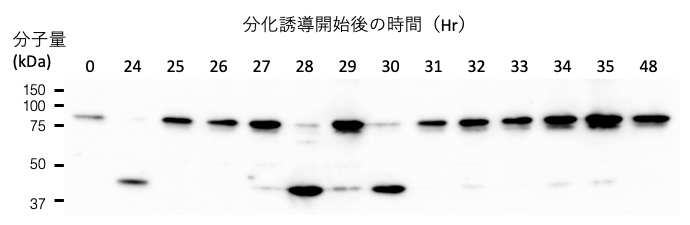
図1 分化誘導された筋芽細胞の中でSTAT3を切断するカルパイン5
分化用培地を用いて筋芽細胞の分化誘導を開始し、翌日1時間ごとに細胞を回収した。ウエスタンブロット法によりSTAT3の解析を行うと24、28、30時間に切断産物が生じていることが分かった。なお、切断のタイミングは実験ごとに異なる。
これらの切断によってSTAT3やc-Mycの全長タンパク質が減少しますが、そのたびに新たなタンパク質合成によってタンパク質の量は元のレベルに復帰しました(図1)。このように間欠泉のような切断現象はタンパク質分解の様式としては珍しい例です。切断により、STAT3やc-Mycは一時的に無力化される状態になるか、または切断によって性質を変えられている可能性があります。
カルパインの一つ、カルパイン5ができるだけ作られなくなるように遺伝子ノックダウンした筋芽細胞中ではSTAT3やc-Mycの切断が抑制されました。このことからカルパイン5が分化過程にある筋芽細胞中で活性化することが示唆されました。このカルパイン5ノックダウン筋芽細胞(ノックダウン細胞)を分化誘導すると、元の細胞と同様に細胞融合を起こして多核細胞を形成しました。ところが、筋肉の収縮運動を起こす筋原線維の構成成分であるミオシンやアクチニンは正常な筋細胞に比べて少量しか作られず(アクチニンの場合5%以下。図2)、筋原線維がきちんと形成されませんでした。また、正常な筋細胞中では多数の核が細胞全体にわたって分布しますが、ノックダウン細胞中の核は1カ所に凝集していました(図3)。さらに、筋分化の進行に伴って減少するはずの活性型STAT3が高いレベルを保ったままでした。人為的に作った活性型STAT3を筋芽細胞中で発現させると、分化誘導条件下でのミオシンやアクチニンの合成が抑えられていました。これらの結果から、カルパイン5の間欠泉様の活性化がこれらのタンパク質レベルを正しく制御するために必要だと考えられます。
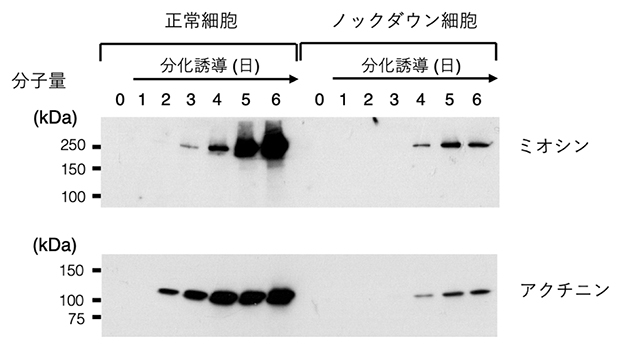
図2 カルパイン5の有無による筋タンパク質の合成
筋芽細胞を分化培地において分化誘導を開始し、24時間ごとに細胞を回収してウエスタンブロット法で解析した。筋細胞の中に形成される筋原線維の主要構成タンパク質であるミオシンやアクチニンは、正常細胞では日を追ってたくさん作られるようになるが、カルパイン5のノックダウン細胞中ではごくわずかしか作られない。
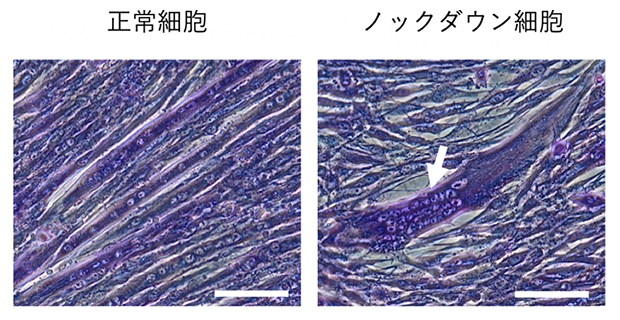
図3 正常細胞には見られないノックダウン細胞中の核の凝集
(左)正常細胞からは線維状の細長い多核細胞(紫色)が作られ、核(白抜け部分)は分散している。
(右)ノックダウン細胞から作られた多核細胞中では核が凝集し(矢印)、細胞が膨らんでいる。スケールバーは100マイクロメートル(μm、1μmは100万分の1メートル)の長さを表す。
今後の期待
筋分化の制御機構はこれまで主に遺伝子発現制御(転写調節因子による制御)の観点から理解されてきましたが、今回の研究で、一過的なタンパク質分解が転写調節に影響を与えることで筋タンパク質量をコントロールするという新たなメカニズムが重層的に働いていることが示唆されました。研究チームはSTAT3、c-Myc以外の転写調節因子もカルパイン5によって切断される可能性があると考えています。このようなタンパク質分解がどのようなメカニズムで遺伝子発現に影響しているかを解明することは重要な研究テーマであり、現在研究チームは、レーザー加工によって作った微小チップ上での細胞培養系開発にも取り組み、細胞分化解析の効率化も目指しています。
カルパイン5の活性化を起こす細胞内の条件を特定し、細胞内カルシウムの動きを詳しく明らかにしていけば、カルパイン5の働きを増強することによって筋肉作りを促進する薬剤の開発につなげられる可能性があります。また、カルパイン5の遺伝子異常は血管新生炎症性硝子体網膜症の原因となることが近年明らかになっています。カルパイン5の基質となるタンパク質の同定をさらに進め、切断箇所のアミノ酸配列を基にしたカルパイン5の阻害剤を開発すればカルパイン5が原因となる疾病の治療薬を開発することにも役立つでしょう。
カルパインの研究では日本の研究者が国際的に大きな貢献をしてきました。しかし、カルパインが健康な生体内で果たす役割のほとんどは不明です。本研究がカルパインに関する研究の新展開の一つのきっかけになることも期待されます。
補足説明
1.カルパイン
システインというアミノ酸が酵素分子の活性中心に位置し、酵素活性がカルシウムに依存するタンパク質分解酵素。ヒト、マウスにはそれぞれ15種類存在する。
2.骨格筋
体を構成する3種の筋肉(骨格筋、平滑筋、心筋)の一つ。手足や体幹の骨に付いて骨格を動かし、個体の運動機能を担う。
3.血管新生炎症性硝子体網膜症
炎症性の目の病気で、網膜の損傷や剝離、硝子体の出血などが起こり、やがては失明に至る。カルパイン5が異常に活性化することによって発症することが遺伝学的解析により示されている。
4.多核の骨格筋細胞になる過程
ヒト成人の骨格筋は筋繊維細胞が束状になったもので、個々の筋繊維細胞は数十から数百個の核を含む多核細胞である。この多核細胞は単核の筋芽細胞(骨格筋前駆細胞)が細胞融合を起こすことによって作られる。
5.遺伝子ノックダウン
遺伝子の転写や翻訳(mRNAを基にしてタンパク質を作る)の段階を抑制、阻害する操作のこと。遺伝子ノックアウトとは異なり、遺伝子の機能を減弱させるものの完全にはなくさない。本研究ではカルパイン5 mRNAを分解させる効果を持つRNAが筋芽細胞中で作られるように遺伝子操作を行った。
6.筋原線維
筋細胞中にあるタンパク質から成る線維状構造。骨格筋では筋細胞の長軸方向に沿って長い線維状構造が形成され、その中でミオシンの重合体とアクチンの重合体の間の滑り運動が起こって線維が縮むことにより筋肉が収縮する。
7.転写調節因子
細胞の核内にあってDNAの特定の塩基配列を認識して結合し、転写(DNAを基にしてmRNAを作る)を促進または抑制する一群のタンパク質。STAT3やc-Mycは、細胞増殖を増進させる働きのあるタンパク質をコードする遺伝子の転写を促進するが、一方でSTAT3の働きは筋分化の進行にとっても必要と考えられている。
8.小胞体
真核生物を構成する細胞内に存在する袋状の構造。細胞外に分泌されるタンパク質や細胞内の膜構造に結合するタンパク質はこの中で作られる。また、細胞質ゾルに比べて数百から千倍程度高い濃度のカルシウムを貯蔵し、それを小胞体外に放出することで細胞内にあるさまざまなカルシウム依存性タンパク質の機能を制御する。
9.小胞体ストレス
細胞に生じるストレスの一種。小胞体内で作られるタンパク質はそれぞれの分子が正しい立体構造を持つ必要があるが、立体構造形成不全を起こしたタンパク質が蓄積した状態を小胞体ストレスと呼ぶ。小胞体ストレスの原因はさまざまあるが、小胞体内カルシウム濃度の低下は代表的な原因の一つである。
10.ウエスタンブロット法
タンパク質試料(精製したタンパク質や細胞を破砕して抽出したタンパク質の混合物など)を膜に固定化したウエスタン・ブロットと呼ばれるものを作製し、試料に含まれる特定のタンパク質の大きさを測ったり、量を測定したりする方法。ブロットは、あらかじめ試料をゲル内で電気泳動し、タンパク質をサイズごとに分離した後で作製する。特定タンパク質の検出、定量には特異抗体が用いられる。
研究支援
本研究は、日本学術振興会(JSPS)基盤研究(C)「筋分化過程に必須な小胞体カルシウム枯渇の分子基盤と細胞内情報伝達における役割(研究代表者:森島信裕)」の助成を受けて行われました。
原論文情報
Nobuhiro Morishima and Yoshihiro Ito, “Calpain-5 regulates muscle-specific protein expression and nuclear positioning during myoblast differentiation”, Journal of Biological Chemistry, 10.1016/j.jbc.2024.107842
発表者
理化学研究所
開拓研究本部 伊藤ナノ医工学研究室(研究当時)
客員研究員(研究当時)森島 信裕(モリシマ・ノブヒロ)
(現 光量子工学研究センター 先端レーザー加工研究チーム 客員研究員)
主任研究員(研究当時)伊藤 嘉浩(イトウ・ヨシヒロ)
(現 光量子工学研究センター 先端レーザー加工研究チーム 客員主管研究員)
報道担当
理化学研究所 広報室 報道担当

