
2022-04-13 理化学研究所
理化学研究所(理研)開拓研究本部眞貝細胞記憶研究室の福田渓研究員(研究当時)、眞貝洋一主任研究員らの共同研究グループは、男性の生殖細胞においてレトロエレメント[1]が制御される仕組みを明らかにしました。
本研究成果は、生殖細胞のエピゲノム[2]形成機構や男性不妊の理解につながると期待できます。
生殖細胞におけるレトロエレメントの異常な活性化は、不妊や次世代の疾患リスクを増大させると考えられていますが、その制御機構は分かっていませんでした。
今回、共同研究グループは、ヒト生殖細胞においてレトロエレメントの抑制に関わるDNAメチル化[3]をゲノムワイドに解析しました。その結果、生殖細胞のもとになる始原生殖細胞[4]では、DNA配列依存的にレトロエレメントがメチル化されるのに対し、精子分化過程では、挿入されたゲノム領域の環境によりメチル化されるか否かが決まることを突き止めました。さらに、精子分化過程でのレトロエレメントのメチル化効率は個人間で異なり、レトロエレメントが精子エピゲノムの個人間多様性を創出することを見いだしました。これらの結果により、男性の生殖細胞におけるレトロエレメントのメチル化制御機構の一端が明らかになりました。
本研究は、オンライン科学雑誌『eLife』(3月22日付)に掲載されました。

男性生殖細胞におけるレトロエレメントのメチル化制御
背景
ヒトゲノムの約半分は、動く遺伝子とも呼ばれるトランスポゾン[5]由来の配列で占められており、その多くはRNAを介して自身のコピーを増幅するレトロエレメントです。レトロエレメントの活性化はゲノムの不安定化を引き起こすだけでなく、がんや精神疾患などさまざまな疾患と関連します。
体細胞ではDNAメチル化によりレトロエレメントの転写が抑制されていますが、この体細胞におけるDNAメチル化は、生殖細胞のもととなる始原生殖細胞ではほぼ完全に失われています。その後生殖細胞形成過程で、精子あるいは卵子特異的なDNAメチル化パターンが形成されます。生殖細胞におけるレトロエレメントの活性化は不妊と関連しますが、DNAメチル化が大きく変化する生殖細胞形成過程で、どのようにレトロエレメントが制御されているのかは分かっていませんでした。
研究手法と成果
共同研究グループは、公開されているヒト精子形成過程の全ゲノムDNAメチル化データを再解析し、精子形成過程におけるレトロエレメントのDNAメチル化の動態を調べました。生殖細胞の源である始原生殖細胞では全ゲノムが脱メチル化しますが、SVA[6]など一部のレトロエレメントではメチル化が維持されていることが分かりました(図1)。
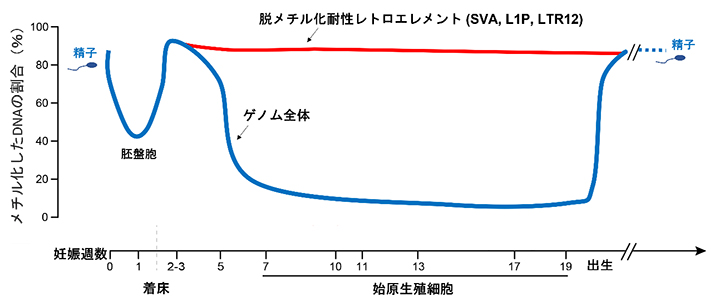
図1 ヒト生殖細胞形成過程のDNAメチル化動態
受精後、胚盤胞になるまでは脱メチル化が進み、その後再メチル化される。生殖細胞系列の細胞は始原生殖細胞に分化するまでに全ゲノムが脱メチル化を受け、精子あるいは卵子への分化過程で再メチル化される(青線)。ただし、霊長類特異的なレトロエレメントのSVA、L1P、LTR12の一部のコピーは、始原生殖細胞における脱メチル化を免れる(赤線)。
そこで、これらのメチル化を維持しているレトロエレメントと関連するDNA配列を探索したところ、DNA結合タンパク質KRAB-ZFP[7](ZNF257やZNF28など)が認識する特定のDNA配列(結合モチーフ)が同定されました。一方、転移活性を示すレトロエレメントではこのKRAB-ZFPの結合モチーフが失われており、始原生殖細胞において脱メチル化されていることが分かりました。このことは、レトロエレメントが宿主の抑制機構を免れるように進化してきたことを示唆しています(図2)。
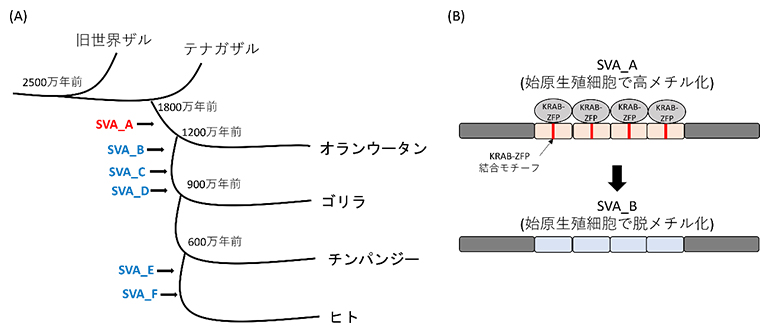
図2 SVAの構造進化と始原生殖細胞でのメチル化状態
(A)霊長類特異的レトロエレメントであるSVAはテナガザルと分岐した後に出現し、進化の過程でコピー数を増やしてきた。宿主の抑制を免れるようにDNA配列が変化したコピーが増幅することで、SVA_A→SVA_B→SVA_C…のように、進化の過程で新たなタイプのSVAが出現してきたと考えられている。今回、最も古いタイプのSVAであるSVA_A(赤文字)のみ、始原生殖細胞で高メチル化されることが分かった。
(B)SVA_Aには、始原生殖細胞で特に高発現するKRAB-ZFP(ZNF257/28)の結合モチーフが複数存在する。一方、始原生殖細胞で脱メチル化する新しいタイプのSVAには、これらのKRAB-ZFP結合モチーフが見られなかった。KRAB-ZFPはDNAメチル化酵素などの転写抑制因子を呼び込むため、新しいタイプのSVAは、KRAB-ZFP結合モチーフを失うことで始原生殖細胞におけるメチル化を免れたと推測される。
次に、始原生殖細胞で脱メチル化されていたレトロエレメントが精子形成過程で再メチル化されるか調べたところ、精子形成過程で転写される領域に転写方向と逆向きに挿入されているレトロエレメントはメチル化されていることが分かりました(図3)。精子形成過程でメチル化されるレトロエレメントとメチル化されないレトロエレメントでDNA配列に大きな違いが見られなかったことから、DNA配列を標的とする機構ではなく、転写産物を標的とする機構によりレトロエレメントはメチル化されると推測しました。さらに、この転写と関連したメチル化の効率は個人差があり、精子におけるレトロエレメントのメチル化の程度が低い個人と高い個人が存在することが分かりました(図3)。
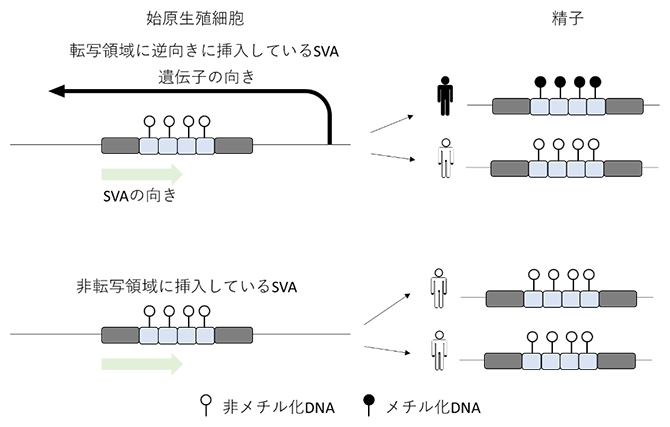
図3 精子形成過程におけるSVAのメチル化動態
上段:始原生殖細胞において脱メチル化を受けたSVAのうち、遺伝子の内部に、遺伝子と逆方向に挿入されたSVAは精子形成過程でメチル化される傾向にある。しかし、その効率は個人間で異なり、精子におけるエピゲノム多様性を形成する。
下段:非転写領域に挿入しているSVAはメチル化されない。
本研究結果から、レトロエレメントは生殖細胞の発生段階で異なる機構によりメチル化されることが明らかになりました。さらに、生殖細胞においてレトロエレメントの抑制状態は個人間で異なることが示唆されました。
今後の期待
本研究では、ヒトの生殖細胞においてどのレトロエレメントがメチル化の標的になるかを明らかにするとともに、精子におけるレトロエレメントのメチル化状態に個人間で多様性があることを突き止めました。
レトロエレメントの活性化は不妊やゲノム異常を引き起こす可能性があることから、本研究成果は男性不妊や遺伝病の形成機構の理解に貢献すると期待できます。また、今後レトロエレメントの抑制状態の個人間の違いの背景にある遺伝因子や環境因子を明らかにできれば、生殖細胞におけるレトロエレメント抑制機構のさらなる解明につながることも期待できます。
補足説明
1.レトロエレメント
転写により生じたRNAから逆転写酵素で自身のコピーを作り、元々存在していたゲノム領域とは異なる領域に挿入することで、コピー数を増幅させる利己的なDNA因子。レトロエレメントの活性化は、がん、不妊、精神疾患などさまざまな病気と関連する。
2.エピゲノム
ゲノムDNAやDNAが巻き付いているヒストンタンパク質に見られる化学修飾の総体。エピゲノムの異常は、がんをはじめとするさまざまな疾患を引き起こすことが知られている。
3.DNAメチル化
哺乳類のDNAメチル化では、主にシトシン・グアニン(CG)配列のCがメチル基で修飾される。体細胞では大部分のCG配列がメチル化しているが、遺伝子発現調節領域ではしばしば低メチル化状態にあり、遺伝子発現調節領域におけるDNAメチル化は基本的に転写抑制と関連している。
4.始原生殖細胞
全ての生殖細胞の源となる細胞種で、胎生期に一時的に出現する。雌では卵原細胞・卵母細胞を経て卵子に分化し、雄では精原細胞・精母細胞・精細胞を経て精子に分化する。始原生殖細胞では体細胞のエピゲノム情報をリセットするため、DNAメチル化の脱メチル化をはじめ、さまざまなエピゲノムのリプログラミングが生じる。
5.トランスポゾン
細胞内でゲノム上の位置を移動(転移)するDNA配列のことで、転移因子ともいう。Cut-And-Paste式で転移するDNA型とCopy-And-Paste式で転移するRNA型の2種類がある。トランスポゾンの転移は宿主に障害をもたらすため、「利己的因子」あるいは「有害因子」といわれてきたが、近年、進化の過程で機能を獲得し、宿主の生存に必須となったトランスポゾンも明らかになっている。
6.SVA
霊長類特異的レトロエレメントで、さまざまな反復配列が融合している。ヒトゲノム上には約2,700コピー存在する。SVA_E/Fはヒト特異的なSVAタイプであり、転移活性を保持している。SVAの挿入は近傍遺伝子の発現やスプライシングパターンに異常を引き起こし、福山型筋ジストロフィーやパーキンソン病などさまざまな疾患と関連することが報告されている。SVAはSINE-VNTR-Aluの略。
7.KRAB-ZFP
KRABドメインのあるC2H2型DNA結合タンパク質で、ヒトゲノムに350種類程度ある。KRAB-ZFPの多くがレトロエレメントに結合し、DNAメチル化酵素などの転写抑制因子をレトロエレメントに呼び込む。KRAB-ZFPとレトロエレメントは互いに競合的に進化してきたと考えられている。KRAB-ZFPはThe Krüpel-associated box zinc finger proteinの略。
共同研究グループ
理化学研究所 開拓研究本部 眞貝細胞記憶研究室
研究員(研究当時) 福田 渓(ふくだ けい)
主任研究員 眞貝 洋一(しんかい よういち)
東京大学 定量生命科学研究所
教授 岡田 由紀(おかだ ゆき)
助教(研究当時) 牧野 吉倫(まきの よしのり)
東京歯科大学
非常勤講師 兼子 智(かねこ さとる)
原論文情報
Kei Fukuda, Yoshinori Makino, Satoru Kaneko, Chikako Shimura, Yuki Okada, Kenji Ichiyanagi, Yoichi Shinkai, “Potential role of KRAB-ZFP binding and transcriptional states on DNA methylation of retroelements in human male germ cells”, eLife, 10.7554/eLife.76822
発表者
理化学研究所
開拓研究本部 眞貝細胞記憶研究室
研究員(研究当時) 福田 渓(ふくだ けい)
主任研究員 眞貝 洋一(しんかい よういち)
報道担当
理化学研究所 広報室 報道担当

