
2022-06-18 理化学研究所,奈良先端科学技術大学院大学
理化学研究所(理研)環境資源科学研究センター細胞機能研究チームの柴田美智太郎研究員、デービッド・ファベロ研究員、杉本慶子チームリーダー、奈良先端科学技術大学院大学の細川陽一郎教授、竹林竜大学院生(研究当時)らの共同研究チームは、栄養が過剰に存在する環境において、シロイヌナズナ[1]が根毛の成長を強く抑制することを発見し、その仕組みを明らかにしました。
本研究成果は、栄養飢餓だけでなく栄養過剰な条件にも適応した植物を作出する技術へと発展していくものと期待できます。
根毛とは、植物の根の表面に存在する毛のような組織であり、根の表面積を大きくすることで、土壌から水や栄養素を効率的に吸収する機能があります。植物は根毛の長さを調節することで、土壌から吸収する栄養の量を制御すると考えられていますが、その詳しい分子機構は明らかではありませんでした。
今回、共同研究チームは、モデル植物[1]であるシロイヌナズナを用いて、栄養が豊富に存在する条件下で根毛の応答を解析しました。そして、通常の生育培地の栄養をたった2倍に(今回のコントロールに対しては4倍)するだけで、シロイヌナズナが根毛をほとんど形成しないことを発見しました。さらに、根毛の成長抑制因子である転写因子[2]GTL1とDF1の二重欠損変異株では、根毛の成長抑制がうまく機能せずに異常な根毛が作られることが分かりました。これらの結果は、シロイヌナズナは栄養が必要以上に存在する条件では、GTL1およびDF1の機能を介して、必要のない根毛の成長を抑制することを示しています。
本研究は科学雑誌『New Phytologist』のオンライン版(6月17日付)に掲載されました。
背景
植物は、自身の成長に必要な栄養素や水分を根から吸収します。根毛は植物の根の表面に存在する毛のような組織であり、根の表面積を大きくすることで土壌から効率的に栄養を吸収します。さらに、植物は根毛の長さを調節することで、土壌から吸収する栄養の量を制御しています。例えば、土壌中に栄養素が少ないときは根毛の成長を促し、不足する栄養素の吸収量を高めます。特に、リン酸や鉄分の欠乏に対する応答が著しいことから、これまではこれら「特定」の栄養素の「飢餓」条件下で、根毛の成長が「促進」される仕組みが精力的に研究されてきました。
一方、近年では安易に入手できる化成肥料のために、時として植物は過剰な栄養条件にさらされる場合があります。しかし、「複数」の栄養素が「過剰」に存在するときの植物の応答に関する知見は多くありません。そこで、共同研究チームは根毛の応答に注目して、過剰量の栄養にさらされたときの植物の応答について研究を行いました。
研究手法と成果
植物研究では、通常の生育条件(栄養条件)としてムラシゲースクーグ培地(MS培地)[3]が広く用いられています。今回、共同研究チームは、MS培地を半分に希釈した培地(1/2xMS)をコントロールとして、2倍濃度にした培地(2xMS)を栄養過剰条件として用意し、それぞれの培地でシロイヌナズナを生育させました。野生株のシロイヌナズナは、1/2xMS培地では通常通り根毛を形成し、2xMS培地ではほとんど根毛を形成しないことが分かりました(図1左)。
そこで、この根毛の成長抑制に強く関わる因子を明らかにするために、これまで知られていた根毛の成長抑制因子の変異株を2xMS培地で生育させて、その根毛の応答を調べました。すると、根毛の成長抑制因子である転写因子GTL1とDF1の二重欠損変異株(gtl1 df1変異株)では、通常の根毛とは異なるバルーン状の根毛を形成することを発見しました(図1右)。このバルーン状の根毛の物理的性質を原子間力顕微鏡(AFM)[4]で測定したところ、野生株の根毛と比較して「固さ」を失った脆弱な構造であることが分かりました。

図1 生育培地の違いによるシロイヌナズナの根毛の変化
左:シロイヌナズナの野生株は、1/2xMS培地で生育させると通常通り根の表面に根毛を形成した。一方、2xMS培地で生育させた場合は、ほとんど根毛を形成しなかった。
右:gtl1df1変異株は、1/2xMS培地で生育させると野生株同様に根の表面に根毛を形成したが、2xMS培地で生育させた場合は、野生株では見られないバルーン状の根毛を形成した。
次に、野生株とgtl1 df1変異株との遺伝子発現解析を行いました。GTL1とDF1は転写因子のため、その変異株では野生株と比較して遺伝子発現に違いがあると予想されます。根毛の成長に関わる遺伝子に絞り込んで解析したところ、gtl1 df1変異株では野生株と比較して転写因子RSL4をコードするRSL4遺伝子の発現量が有意に上昇していることを見いだしました。RSL4は根毛成長のマスターレギュレーター[5]だと考えられており、RSL4タンパク質の量と根毛の長さには強い相関関係があります。従って、gtl1 df1変異株で不完全な根毛が形成されたことは、RSL4遺伝子の発現上昇が原因と考えられます。
また、RSL4遺伝子は別の転写因子RHD6によって直接発現誘導されることが知られています。そこで、RHD6とGTL1との関係をさらに詳しく調べました。まず培養細胞を用いたルシフェラーゼレポーターアッセイ[6]から、RHD6とGTL1が同時に存在する場合は、RSL4遺伝子に対してGTL1による抑制機構が優先されることが明らかになりました。さらに、共免疫沈降解析[7]から、GTL1とRHD6が物理的に相互作用することが示されました。これらの結果は、GTL1がRHD6と複合体を形成し、その機能を阻害することでRSL4遺伝子の発現を抑制することを示唆しています(図2)。
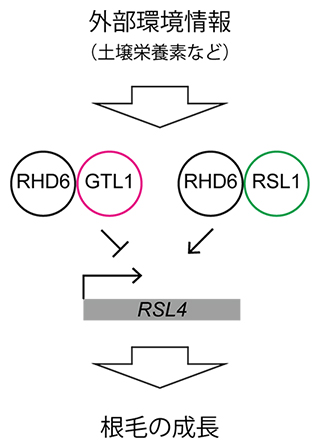
図2 本研究から示唆される外部環境依存的な根毛の成長を制御する機構
外部環境に応じて、RHD6は抑制型の複合体RHD6-GTL1や活性型の複合体RHD6-RSL1へと変化する。その結果、下流のRSL4遺伝子の発現量が調整され、根毛の成長が制御される。
最後に、RSL4遺伝子の発現上昇がgtl1 df1変異株におけるバルーン状の根毛の原因であることを示すために、rsl4 gtl1 df1三重変異株を作製し、2xMS培地での表現型を観察しました。まず図1右側でも示したように、gtl1 df1変異株は2xMS培地で生育させるとバルーン状の根毛を形成します(図3左)。一方、rsl4 gtl1 df1三重変異株では、gtl1 df1変異株で観察されるバルーン状の根毛は観察されませんでした(図3中)。さらに、35Sプロモーターを用いてRSL4遺伝子の過剰発現体を観察したところ、2xMS培地でgtl1 df1変異株のようなバルーン状の根毛が形成されることが分かりました(図3右)。これらの結果は、本来GTL1とDF1によって抑制されるはずのRSL4遺伝子が、gtl1 df1変異株では適切に発現抑制されず、その結果として異常な根毛形成につながったことを示唆しています。
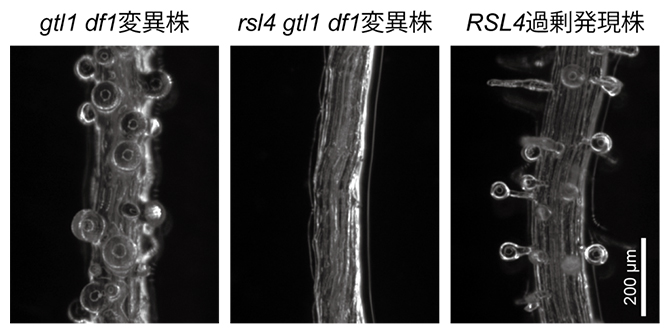
図3 RSL4遺伝子の過剰発現によって引き起こされる根毛形成異常
gtl1 df1変異株は2xMS培地で生育させるとバルーン状の根毛を形成したが、rsl4 gtl1 df1三重変異株はバルーン状の根毛を形成しなかった。一方で、RSL4過剰発現株を2xMS培地で生育させると、gtl1 df1変異株のようなバルーン状の根毛が形成された。
今後の期待
本研究から、過剰に栄養素が存在する条件下で植物は根毛の成長が著しく抑制されること、その抑制には転写因子GTL1とDF1によるRSL4遺伝子の発現抑制が重要であることが明らかになりました。この応答は、栄養が豊富に存在する環境において不必要な根毛を作らない「省エネ」のためのメカニズムであると考えられます。
本研究は実験室環境でのみの検証でしたが、過剰に施肥された農地など、植物が過剰量の栄養素にさらされることは現実に存在します。今後の期待としては、野外や農地での実験を通して、栄養飢餓条件だけでなく過剰条件にも適応したより生育できる幅の広い植物を作出する技術へつなげていくことが考えられます。
また、本研究成果は、国際連合が2016年に定めた17項目の「持続可能な開発目標(SDGs)[8]」のうち「2.飢餓をゼロに」と「15.陸の豊かさも守ろう」に大きく貢献するものです。
補足説明
1.シロイヌナズナ、モデル植物
アブラナ科の植物の一つで、「ぺんぺん草」で知られるナズナの近縁種である。持っている遺伝子の総量が比較的少なく、発芽から開花し種子が採れるまでの時間が比較的短いことから、被子植物の代表として世界中で研究対象になっている。2000年に植物としては初めて全遺伝子の解読が終了し、特に分子生物学分野で多用されている。普遍的な生命現象の解明に用いられる代表的な生物をモデル生物と呼ぶが、シロイヌナズナはモデル植物となっている。
2.転写因子
DNAに結合するタンパク質。遺伝子の発現量を調節する機能があり、発生や成長に重要な役割を担う。
3.ムラシゲ-スクーグ培地(MS培地)
植物の生育に必要な栄養素が含まれた培地。植物栽培用培地として広く使用されている。シロイヌナズナの場合は、半分の濃度に希釈した1/2xMSで使用されることが多い。
4.原子間力顕微鏡(AFM)
探針で物質の固さを測定する技術。レーザーを用いて探針のたわみを検出することで、ナノメートル(10億分の1メートル)~マイクロメートル(100万分の1メートル)スケールの解像度で対象の物理的性質を測定することができる。AFMはAtomic Force Microscopeの略。
5.マスターレギュレーター
ある現象に関与する遺伝子発現制御の最上位に位置する制御因子のこと。転写因子RSL4は根毛の成長に必要な遺伝子群(細胞壁の合成など)を直接発現制御し、根毛の成長を直接的に調整する。ゆえに、RSL4は根毛成長のマスターレギュレーターである。
6.ルシフェラーゼレポーターアッセイ
転写因子とプロモーターとの間の相互作用を検出する方法。プロモーターとは遺伝子の上流領域を指し、この領域に転写因子などのタンパク質が結合することで遺伝子発現が調節されると考えられている。プロモーター領域下流にルシフェラーゼ遺伝子を融合させた遺伝子(レポーター)と転写因子(エフェクター)を同時に発現させ、ルシフェラーゼに由来する発光強度を測定する。発光強度がコントロール(通常は空ベクター)に対して変化が見られた場合、その転写因子とプロモーター領域との間に相互作用が見られたと見なす。本研究では、シロイヌナズナ培養細胞MM2dを用いた一過的な形質転換体で行った。
7.共免疫沈降解析
タンパク質間の相互作用を検出する実験手法。まず、標的とするタンパク質を特異的な抗体で捕捉・精製(免疫沈降)する。この際、標的タンパク質と相互作用するタンパク質も同時に精製(共免疫沈降)される。本研究では、FLAGとHAというアフィニティタグを用い、ウェスタンブロットにより相互作用するタンパク質を検出した。
8.持続可能な開発目標(SDGs)
2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標。持続可能な世界を実現するための17のゴール、169のターゲットから構成され、発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本も積極的に取り組んでいる(外務省ホームページから一部改変して転載)。
共同研究チーム
理化学研究所 環境資源科学研究センター 細胞機能研究チーム
チームリーダー 杉本 慶子(すぎもと けいこ)
研究員 柴田 美智太郎(しばた みちたろう)
研究員 デービッド・ファベロ(David S.Favero)
テクニカルスタッフⅡ 竹林 有理佳(たけばやし ありか)
テクニカルスタッフⅡ 河村 彩子(かわむら あやこ)
研究員(研究当時) バート・ライメン(Bart Rymen)
(現 ルーヴェンカトリック大学植物研究所(LPI))
奈良先端科学技術大学院大学 先端科学技術研究科 物質創成科学領域
教授 細川 陽一郎(ほそかわ よういちろう)
大学院生(研究当時) 竹林 竜(たけばやし りゅう)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費補助金特別研究員奨励費「根毛細胞の可塑的な伸長成長を支える転写制御ネットワークの解明(特別研究員:柴田美智太郎)」「AHL転写因子を介した植物の器官成長を司る転写制御ネットワークの解明(受入研究者:杉本慶子)」、同若手研究「転写因子GTL1の機能調節機構の解明(研究代表者:柴田美智太郎)」、同学術変革領域研究(A)「不均一環境変動に対する植物のレジリエンスを支える多層的情報統御の分子機構」の研究課題「植物の環境レジリエンスを支える傷害修復機構(研究代表者:杉本慶子)」、同新学術領域(研究領域提案型)「植物の力学的最適化戦略に基づくサステナブル構造システムの基盤創成」の研究課題「顕微技術を駆使した計測と制御による細胞構造のしなやかさの高精度解析(研究代表者:細川陽一郎)」、科学技術振興機構(JST)戦略的創造研究推進事業ACT-X「環境とバイオテクノロジー」の研究課題「根毛と遺伝子制御ネットワークを軸とした植物環境応答機構の解明(研究代表者:柴田美智太郎)」による助成を受けて行われました。
原論文情報
Michitaro Shibata, David S. Favero, Ryu Takebayashi, Arika Takebayashi, Ayako Kawamura, Bart Rymen, Yoichiroh Hosokawa, Keiko Sugimoto, “Trihelix transcription factors GTL1 and DF1 prevent aberrant root hair formation in an excess nutrient condition”, New Phytologist, 10.1111/nph.18255
発表者
理化学研究所
環境資源科学研究センター 細胞機能研究チーム
チームリーダー 杉本 慶子(すぎもと けいこ)
研究員 柴田 美智太郎(しばた みちたろう)
研究員 デービッド・ファベロ(David S.Favero)
奈良先端科学技術大学院大学 先端科学技術研究科 物質創成科学領域
教授 細川 陽一郎(ほそかわ よういちろう)
大学院生(研究当時) 竹林 竜(たけばやし りゅう)
報道担当
理化学研究所 広報室 報道担当
奈良先端科学技術大学院大学 企画・教育部企画総務課渉外企画係
