2024-04-17 理化学研究所,福井県立大学,宮崎大学
理化学研究所(理研)仁科加速器科学研究センター イオン育種研究開発室の阿部 知子 室長、石井 公太郎 協力研究員(研究当時、現 客員研究員)、平野 智也 協力研究員(研究当時、現 客員研究員、宮崎大学 農学部 准教授)、イオン育種研究開発室植物ゲノム進化研究チーム(研究当時)の風間 裕介 チームリーダー(研究当時、現イオン育種研究開発室 客員研究員、福井県立大学 生物資源学部 教授)らの研究チームは、重イオンビーム[1]で誘発するDNA欠失変異の最大値や、染色体構造変化を決めるのはゲノム中に散在する必須遺伝子[2]であることを実証しました。
これまで、重イオンビームが誘発する欠失変異では、ビームの線エネルギー付与(LET)[3]が重要であり、誘発する変異サイズはLETに依存し、LETが大きいほど変異サイズも大きくなると考えられていました。
今回、研究チームはLETが100~290キロ電子ボルト/マイクロメートル(keV/μm)では欠失の大きさに差がないことを明らかにしました。この原因を探るため、シロイヌナズナのゲノムに対して、孫世代に遺伝した欠失と必須遺伝子の位置を、塗り絵のようにして染色体上にマップしたところ、必須遺伝子を含む欠失は次世代に遺伝しにくいことが分かりました。また、確実に遺伝する欠失の大きさは必須遺伝子間の距離よりも小さいことも判明しました。これらのことからゲノム中の必須遺伝子の位置が欠失の大きさの上限を決めることが示唆されました。さらに、染色体構造変化のつなぎ目は必須遺伝子を避けるように生じていることも明らかになりました。本研究成果は、必須遺伝子の分布から欠失の最大値を推定し、適正なLETを選択することにより、重イオンビームを用いた突然変異育種のさらなる効率化に貢献すると考えられます。
本研究は、科学雑誌『Frontiers in Plant Science』オンライン版(4月17日付:日本時間4月17日)に掲載されました。

欠失の大きさに関与する二つの要因(本研究で②が実証された)
背景
電離放射線の一種である重イオンビームは効果的に突然変異を誘発することのできる変異原として、植物や微生物の品種改良に用いられています。研究チームはこれまでに花の新品種、清酒酵母やパン酵母など41の製品を市場に提供しました。理研RIビームファクトリーでは、線エネルギー付与(LET)を幅広く選択できるため、植物種や目的とする遺伝子によってLETを調整するテーラーメイド品種改良法を開発しています。
選抜した変異体を用いたこれまでの研究では、23~30keV/μm、101~124keV/μmと290keV/μmの重イオンビームにより誘発される遺伝子領域の変異の比較から、線エネルギー付与の高い重イオンビームほど大規模な変異を誘発しやすいと予想していました。今回は、重イオンビームの線エネルギー付与という物理的要因に加え、変異誘発の対象である植物ゲノムの中で、必須遺伝子の分布に注目して解析しました。
研究手法と成果
研究チームは、シロイヌナズナの乾燥種子にそれぞれ100keV/μm、200keV/μm、290keV/μmの3種類のLETの重イオンビームを照射し、照射個体から孫の世代(M3)を育成しました。M3に遺伝した欠失変異をDNAマイクロアレイ[4]や次世代シーケンサー[5]により検出しました。それぞれの欠失の大きさを調べると、欠失の大きさは3種類のLET間で差がありませんでした(図1A)。このことから欠失の大きさはLETの大きさによって単調に増加するわけではなく、上限が存在することが示唆されました。ホモ接合型[6]でも遺伝する欠失(ホモ欠失)と、ホモ接合型では生存できなくなるためヘテロ接合型[6]でのみ遺伝する欠失(ヘテロ欠失)に分けて大きさを比較すると、ヘテロ欠失の方が大きいことが分かりました(図1B)。これは大きい欠失ほどホモ接合型で次世代に遺伝しづらいことを示しています。また、ゲノム中の必須遺伝子間の距離の中央値よりもホモ欠失の大きさの中央値の方が小さいことが分かりました(図1B)。従って、シロイヌナズナゲノム中の必須遺伝子が変異の次世代への遺伝を抑制して、ホモ欠失の大きさの上限を規定すると考えられました。
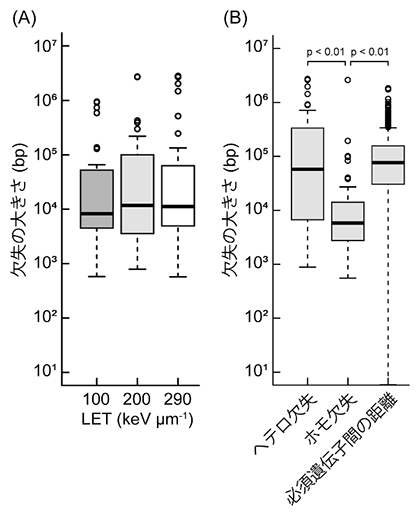
図1 シロイヌナズナ欠失変異の大きさ
(A)各LETで誘発した欠失の大きさを示す。(B)ヘテロ欠失、ホモ欠失に分類した欠失の大きさおよび必須遺伝子間の距離から推定したホモ欠失の上限値を示す。
そこで、シロイヌナズナゲノム上の94個のホモ欠失と55個のヘテロ欠失の位置と、811個の必須遺伝子の位置を比較したところ、ホモ欠失には必須遺伝子が一つも含まれなかったのに対し、ヘテロ欠失は必須遺伝子を含んでいました(図2)。このことから欠失の遺伝には必須遺伝子が重要な役割を持つことが分かりました。
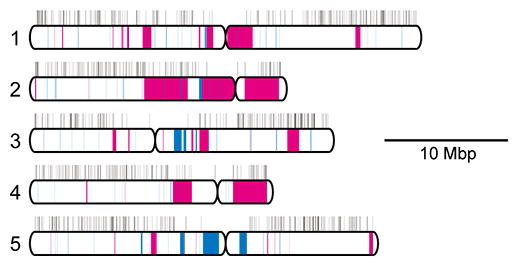
図2 シロイヌナズナ染色体への欠失変異のマップ
シロイヌナズナの五つの染色体をおのおの白い棒で示す。染色体上部の黒い縦線は必須遺伝子の位置を示す。照射個体から検出されたヘテロ欠失の位置を赤、ホモ欠失の位置を青でそれぞれ示す。染色体左側の数字は染色体番号を示す。Mbp(メガベースペア)はDNAの長さの単位で1Mbpは100万塩基対を示す。
研究チームは、研究チームが開発した変異解析パイプラインAMAP注)を用いて、重イオンビームで誘発されて生じた染色体の構造変化の位置を、本研究を含めて340カ所特定しています。これらと他のグループにより明らかにされた92カ所を合わせて、必須遺伝子の位置と照らし合わせました。その結果、染色体の構造変化は必須遺伝子を避けるように生じることが分かりました(図3)。
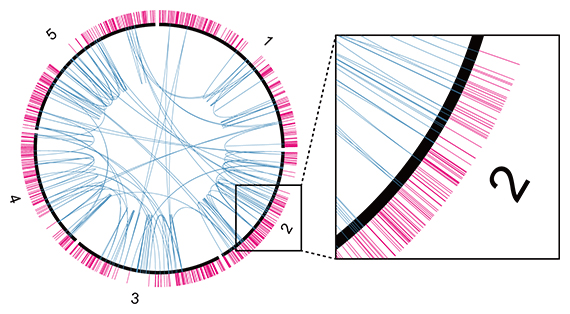
図3 必須遺伝子と染色体構造変化の位置関係
シロイヌナズナの染色体を黒い円で表し、染色体番号を数字で表す。染色体外側のマゼンタの先は必須遺伝子の位置を表す。染色体内側の青の弧は染色体構造変化の生じた位置を示す。染色体構造変化は変異可能な遺伝子の内部や遺伝子外領域では生じていたが、必須遺伝子内部には生じていなかった。
注)Ishii et al., AMAP: A pipeline for whole-genome mutation detection in Arabidopsis thaliana, Genes Genet. Syst. 91(4):229-233 (2016)
今後の期待
研究チームは、「テーラーメイド変異誘発技術」、すなわち、LETを調節することで、ユーザーのさまざまな要望、例えば、とにかく高率で変異体を得たい、目的の遺伝子のみ小さく破壊したい、あるいはゲノムに大規模な変異を起こしてこれまでにない変異体を得たい、などの要望に応える技術の開発に取り組んでいます。本研究で、植物ゲノム中の必須遺伝子の分布が欠失の大きさの上限を決め、欠失サイズの誘発に適正なLETがあるという新しい知見を得ました。これにより、育種の対象となる植物種の必須遺伝子の情報から、得られる欠失サイズを予測し、その植物種に対する変異誘発に適したLETを選択できるなど、より効率的な対応が可能になると考えられます。また、今後は必須遺伝子に続く生物学的側面として、生物が持つ複数のDNA修復経路に注目し、変異誘発に有効な、DNAの間違い(エラー)を起こしやすい修復経路の応答がLETによって異なるかどうかを調査したいと考えています。
また、本研究では染色体の構造変化も必須遺伝子を避けるように生じることが分かりました。染色体の構造変化は生物の進化の過程で起こります。しかし、それが全くランダムなのか、何かしらの法則性があるのかは解明されていません。本研究の結果、必須遺伝子上では構造変化が起こらないことが実証されました。今後、染色体構造変化のデータを蓄積することで、染色体レベルでの生物進化の道筋の理解に寄与することが期待できます。
補足説明
1.重イオンビーム
原子から電子をはぎ取って作られたイオンの中で、ヘリウムイオンより重いイオンを重イオンと呼ぶ。速度と方向がそろった重イオンの粒子束を重イオンビームと呼ぶ。
2.必須遺伝子
ゲノム中にある遺伝子のうち、生育に欠かせない遺伝子や次世代を生産するのに必要な遺伝子のこと。
3.線エネルギー付与(LET)
エネルギーを持った粒子が物質中を通過する際、単位長さ当たりにイオンが失うエネルギー量のこと。単位はキロ電子ボルト/マイクロメートル(keV/μm)などで表され、イオンの種類や速さによって異なる。イオンが失ったエネルギーは通過した物質に局所的に与えられるので、一般に線エネルギー付与が大きいと、生体に与える影響は大きい。LETはlinear energy transferの略。
4.DNAマイクロアレイ
数万から数十万に区切られたスライドガラス上にプローブと呼ばれるDNAの部分配列を高密度に配置し固定したもの。本研究ではシロイヌナズナ野生株のゲノムDNA配列の一部をプローブとして用いた。DNAマイクロアレイに重イオンビーム照射個体のゲノムDNAを反応させると、相補的な配列を持つプローブと結合する。しかし欠失領域の配列に対応するプローブには結合しないため、これを利用して欠失領域を検出することができる。
5.次世代シーケンサー
DNA配列を解読する機械。数千から数百万ものDNA分子を同時に配列決定することが可能であり、シロイヌナズナが持つほぼ全てのDNAのセット(ゲノム)を解読できる。
6.ホモ接合型、ヘテロ接合型
シロイヌナズナの細胞核には対となる二つの染色体が5セット存在する。重イオンビーム照射によりある一つの染色体に変異が誘発され、それが自殖により遺伝する場合を考えると、減数分裂により花粉の中の精細胞には変異した染色体か、変異を持たない対となる染色体のどちらかが含まれる。同様に胚珠(はいしゅ)の中の卵細胞にも変異した染色体か、変異を持たない対となる染色体のどちらかが含まれる。このような植物体が自殖したとき、後代の細胞の対となる染色体がどちらも変異を持つ場合、またはどちらも変異を持たない場合をホモ接合型といい、片方のみ変異を持つ場合をヘテロ接合型という。
研究チーム
理化学研究所 仁科加速器科学センター
イオン育種研究開発室
室長 阿部 知子(アベ・トモコ)
協力研究員(研究当時)石井 公太郎(イシイ・コウタロウ)
(現 客員研究員、量子科学技術研究開発機構放射線医学研究所 計測・線量評価部 主任研究員)
協力研究員(研究当時)平野 智也(ヒラノ・トモナリ)
(現 客員研究員、宮崎大学 農学部 准教授)
イオン育種研究開発室 植物ゲノム進化研究チーム(研究当時)
チームリーダー(研究当時)風間 裕介(カザマ・ユウスケ)
(現 イオン育種研究開発室 客員研究員、福井県立大学 生物資源学部 教授)
研究支援
本研究は、内閣府戦略的イノベーション創造プログラム(SIP)次世代農林水産業創造技術「戦略的オミクス育種技術体系の構築(研究代表者:阿部知子)」、重イオンビームにより誘発される染色体再構成を利用した新育種技術の開発事業(福井県)「超高LETイオンビームによる変異誘発の多様性の実証(研究代表者:阿部知子)」、日本学術振興会(JSPS)科学研究費助成事業基盤研究(C)「重イオンビームによる高頻度変異誘発の分子基盤の解明(研究代表者:石井公太郎)」、同基盤研究(B)「染色体微細加工で逆位・転座が植物ゲノムに与える影響を見る(研究代表者:風間裕介)」、同挑戦的研究(萌芽)「植物ゲノムは遺伝子のコピー数の減少に強いのか(研究代表者:風間裕介)」、同国際共同研究加速基金(国際共同研究強化(B))「植物性染色体の誕生と性決定システムの進化を解明する日英共同研究(研究代表者:風間裕介)」、同学術変革領域研究(B)「起源の新しい植物性染色体に性の消滅回避の兆候を見出す(研究代表者:風間裕介)」による助成を受けて行われました。
原論文情報
Kotaro Ishii, Yusuke Kazama, Tomonari Hirano, Jeffrey A. Fawcett, Muneo Sato, Masami Yokota Hirai, Fujiko Sakai, Yuki Shirakawa, Sumie Ohbu and Tomoko Abe, “Genomic view of heavy-ion induced deletions associated with distribution of essential genes in Arabidopsis thaliana”, Frontiers in Plant Science, 10.3389/fpls.2024.1352564
発表者
理化学研究所
仁科加速器科学センター イオン育種研究開発室
室長 阿部 知子(アベ・トモコ)
協力研究員(研究当時)石井 公太郎(イシイ・コウタロウ)
(現 客員研究員)
協力研究員(研究当時)平野 智也(ヒラノ・トモナリ)
(現 客員研究員、宮崎大学 農学部 准教授)
イオン育種研究開発室 植物ゲノム進化研究チーム(研究当時)
チームリーダー(研究当時)風間 裕介(カザマ・ユウスケ)
(現 イオン育種研究開発室 客員研究員、福井県立大学 生物資源学部 教授)
報道担当
理化学研究所 広報室 報道担当
福井県立大学 経営企画部 連携・研究課
宮崎大学 企画総務部総務広報課
の臨床試験を開始~病理医を必要としない、がん診断の新技術~")
