2021-07-27 東京大学
- 発表者
- 平瀬 祥太朗(東京大学大学院農学生命科学研究科 附属水産実験所 助教)
山崎 曜(国立遺伝学研究所 ゲノム・進化研究系 日本学術振興会特別研究員)
西迫 真登(東北大学 大学院農学研究科 附属複合生態フィールド教育研究センター 大学院生:当時)
池田 実(東北大学 大学院農学研究科 附属複合生態フィールド教育研究センター 教授)
原 素之(東北大学 マリンサイエンス復興支援室 教授:当時)
Juha Merilä(香港大学 生態学・生物多様性研究部門 教授)
菊池 潔(東京大学大学院農学生命科学研究科 附属水産実験所 教授)
発表のポイント
- 日本産のアワビが種分化(注1)しつつあることをゲノム解析によって示した。
- 生息水深の違いなどの生態的な要因が、海産無脊椎動物の種分化をスタートさせる。
- 海産無脊椎動物における種分化の仕組みを解明することにつながり、日本産アワビの保全管理についても重要な指針を提示するものである。
発表概要
高級な水産物である大型アワビ。日本ではエゾアワビとクロアワビ、マダカアワビ、メガイアワビが漁獲されています。これらの種は、北アメリカ沿岸に生息していた祖先種が日本列島沿岸に分布拡大することで最近に誕生したと考えられており、遺伝的に非常に近い関係にあるものの、自然界において独自の形態的・生態的特徴を有しています。その一方で、全ての種は実験室下では交配させることができることから、種分化が完了していない可能性がこれまで指摘されていました。本研究は、日本産アワビのゲノム解析を行い、これらの種が交雑を伴いながら、今まさに種分化しようとしていることを明らかにしました。本研究の成果は、海産無脊椎動物における新種誕生のメカニズムの解明へとつながり、また、日本産アワビの保全管理を行う上で重要な情報となります。
発表内容

図1 日本産アワビ3種の形態的・生態的特徴
エゾアワビとクロアワビは亜種の関係にある。地図上の矢印はエゾアワビとクロアワビの境界域を示す。
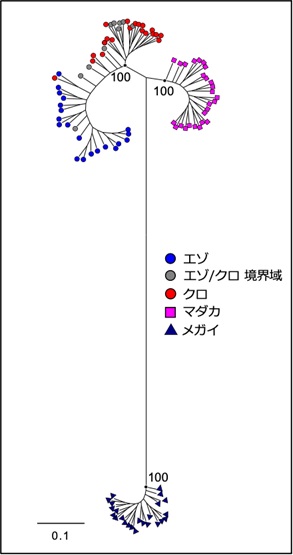
図2 ゲノムワイドSNPマーカーに基づく日本産アワビの分子系統樹
数値はブートストラップ確率を示す。エゾ・クロアワビとマダカアワビ、メガイアワビは、それぞれ高いブートストラップ確率で異なるクレードを形成し、自然界において生殖的隔離が生じていることが示された。
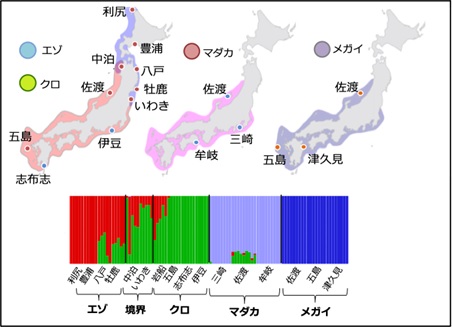
図3 ゲノム解析に用いた日本産アワビの地域集団とクラスタリング分析
横に並ぶ一つ一つのバープロットが各個体におけるゲノムの混ざり合いを示し、色は、各種のゲノムを示す。エゾアワビとクロアワビは亜種の関係にあり、ゲノムが混ざっている。一方で、エゾ・クロアワビ、マダカアワビ、メガイアワビの間ではゲノムの共有がほとんど生じておらず、現在はほとんど交雑がないことが示唆される。一方、佐渡のマダカアワビは、クロアワビのゲノムを一部保有しており、この地域においては種間の生殖的隔離が弱いことが推測される。
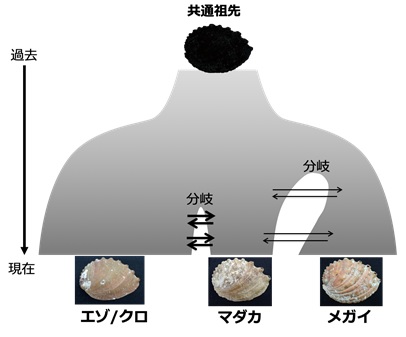
図4 デモグラフィックモデリングで推定された日本産アワビの種分化プロセス
図中の矢印は種間の交雑に由来する遺伝子流動(ゲノムの交換)を示す。メガイアワビがまず分岐し、その次にエゾ・クロアワビとマダカアワビが分岐した。それぞれの種が分岐して以降も、3種は遺伝子流動を伴いながら種分化してきたと推測された。
日本列島を取り囲み、地球表面の約70%を占める海洋には膨大な生物種が存在しています。一方で、海洋においては遺伝子流動が盛んに生じ、生物集団間におけるゲノム変異の蓄積が妨げられ、それに伴う種分化が起こりにくいと考えられています。では、どのようなメカニズムで種分化が始まるのでしょうか?この問いは海洋生物学において重要なトピックであり続けています。特に、海産無脊椎動物では卵と精子が海洋に放出され、遺伝子流動が広範に起こるため、種分化が起こりにくいと考えられています。このような海産無脊椎動物の種分化のプロセスを説明する数少ない実例の一つが、北アメリカ沿岸に生息するアワビ類における配偶子のタンパク質をコードする遺伝子(精子のLysinと卵のVERL)(注2)の変異です。これらのアワビ類では、これらの変異が生じていることで異種間の受精が起こらず、生殖的隔離(注3)が生じていることが示唆されています。では、アワビのような海産無脊椎動物の種分化は、このような配偶子による隔離から始まるのでしょうか?それとも、他の要因が種分化を引き起こすのでしょうか?
この問いに答えるため、平瀬助教らは日本沿岸で漁獲されるエゾアワビ(エゾ)とクロアワビ(クロ)、マダカアワビ(マダカ)、メガイアワビ(メガイ)に着目しました。これらの大型アワビは北アメリカ沿岸に分布していた祖先種が、日本列島に分布拡大することで最近に種分化したと考えられており、遺伝的に非常に近い関係にあるものの、独自の形態的・生態的特徴を有しています(図1)。まず、エゾとクロはHaliotis discusに属する北方系と南方系の亜種(注4)と考えられており、10m以浅の比較的浅い水深帯に生息します。そして、分布域がクロと重複するマダカ(Haliotis madaka)は15m以深、メガイ(Haliotis gigantea)はクロとマダカの中間的な水深に生息する別種とされています。その一方で、全ての種は実験室下では交配させることができ、交雑第一世代も生殖が可能なことが明らかになっています。これらの事実は、日本産アワビにおいては生息水深の違いなどの生態学的な要因によって種の独自性が維持されており、そのような生態的な隔離が、配偶子による隔離に先んじて海産無脊椎動物の種分化を促進する可能性を示しています。そこで、平瀬助教らは、これらの日本産アワビのゲノム解析を行うことで、それらの遺伝的な関係性や種分化のプロセスについて研究しました。
まず、3種のアワビからゲノムワイドに取得したSNP(注5)座の塩基配列を用い、分子系統樹(注6)を作成しました。その結果、高いブートストラップ値(注7)によって支持される3つのクレード(注7)が示され、それぞれがエゾ・クロとマダカ、メガイの種の分類と一致しました。したがって、3種が自然界において生殖的に隔離されていることが遺伝学的に確かめられました(図2)。次に、クラスタリング分析によって個体ごとのゲノム構成を調べたところ、種間の交雑に由来する遺伝子流動はほとんど検出されなかった一方で、新潟の佐渡島のマダカにはエゾ・クロからの遺伝子流動が生じていることが示されました(図3)。したがって、エゾ・クロとマダカ間における生殖的隔離の強さは地域によって異なっており、種分化が今まさに進行していることが明らかになりました。また、種間のアリル頻度パターンから過去の集団動態を推定するデモグラフィックモデリング解析(注8)は、これら3種のアワビが交雑による遺伝子流動を伴いつつ種分化してきた歴史を示しました(図4)。このような遺伝的に近い種群の場合、ゲノム全体における遺伝的分化の程度は小さい一方、生殖的隔離に関与するゲノム領域では分化の程度は大きくなっていると予想されます。このようなゲノム領域を探索した結果、複数の染色体に散在する24のゲノム領域を特定することに成功し、これらの領域に日本産アワビの種分化をもたらした遺伝子が存在すると考えられました。今後、これらのゲノム領域を詳細に研究することで、海産無脊椎動物において新種誕生をもたらす遺伝基盤を発見することができるかもしれません。
エゾとクロ、マダカ、メガイは調理法によって違う味わいと食感を示すといいます。このような“味の多様性”は、本研究で明らかとなった種分化プロセスにおける副産物として生じてきました。本研究の成果は、近年、漁獲量が低下している日本産アワビの保全管理や育種を行う上で重要な知見となります。また、さらなる研究によって新たな種分化モデルの提出につながり、日本産アワビが将来の海洋生物の多様性研究に大きな進展をもたらすものと期待されます。
発表雑誌
- 雑誌名
- Molecular Biology and Evolution
- 論文タイトル
- Genomic evidence for speciation with gene flow in broadcast spawning marine invertebrates
- 著者
- Shotaro Hirase*, Yo Y. Yamasaki, Masashi Sekino, Masato Nishisako, Minoru Ikeda, Motoyuki Hara, Juha Merilä, Kiyoshi Kikuchi
- DOI番号
- 10.1093/molbev/msab194
- 論文URL
- https://doi.org/10.1093/molbev/msab194
問い合わせ先
東京大学大学院農学生命科学研究科 附属水産実験所
助教 平瀬 祥太朗(ひらせ しょうたろう)
E-mail:cashirase<アット>g.ecc.u-tokyo.ac.jp <アット>を@に変えてください。
用語解説
注1 種分化
一つの生物種から、別の新しい種が生まれること。
注2 LysinとVERL遺伝子
アワビの精子が持つタンパク質であるLysinと卵が持つタンパク質であるVitelline Envelope Receptor for Lysin(VERL)の相互作用により受精が起こる。北アメリカのアワビでは、これらのタンパク質を支配する遺伝子が種間で異なっており、別種間の受精を妨げていると考えられている。
注3 生殖的隔離
二つの集団間で交配が起こらないことを指す。同所的に存在する集団間で生殖的隔離が生じていることは、それらが別種であることの証拠となる。生殖的隔離は、生殖行動が妨げられることなどにより配偶子の接合(受精)に至らない接合前隔離と、配偶子の接合は起こるが、雑種の生育ないし繁殖が不可能になる接合後隔離に大きく分けられる。
注4 亜種
同一の種に属する地理的な品種。
注5 SNP
Single Nucleotide Polymorphism(一塩基多型)の略。個体間においてゲノムDNA上の1塩基が異なる現象のことを指す。SNPがある場所のことをSNP“座(locus)”と呼ぶ。
注6 分子系統樹
個体のゲノム配列に基づき、それらの進化的な分岐関係を表した図。本研究では、SNP座における塩基配列を用いた。
注7 クレードとブートストラップ確率
分子系統学において、1つの共通祖先から派生した分類群からなるグループを指す。クレードの信頼性は、各分岐点(ノード)におけるブートストラップ確率によって評価される。
注8 デモグラフィックモデリング
各種が持つアリル頻度のパターンから、それぞれの生物集団の過去の動態(集団のサイズや集団間の遺伝子流動の有無)をシミュレーションベースで推定する手法。

