2022-08-22 理化学研究所,産業技術総合研究所
理化学研究所(理研)バイオリソース研究センター微生物材料開発室の加藤真悟上級研究員、大熊盛也室長、産業技術総合研究所地圏資源環境研究部門地圏微生物研究グループの金子雅紀研究グループ付らの共同研究グループは、好気条件で容易に培養可能な、数百ナノメートル(nm、1nmは10億分の1メートル)サイズの絶対寄生性ナノアーキア(古細菌)[1]のリソース化に成功しました。
本研究成果は、「微生物ダークマター」(機能未知の未培養微生物群の総称)の代表格である、寄生もしくは共生性ナノアーキアのさらなる培養・リソース化への足がかりになるだけでなく、ナノアーキアの謎に包まれた生態学的役割や、初期生命の進化過程の解明にも貢献すると期待できます。
今回、共同研究グループは、国内の酸性温泉から、細胞サイズが数百nmのナノアーキアを培養することに成功しました。このナノアーキアは、宿主となるアーキアの細胞表面に付着し、細胞膜に穴を開けて、そこから細胞質を吸い取るという類例のない増殖方法をとっていることが示唆されました。
共同研究グループは、このナノアーキアをNanobdella aerobiophilaと命名し、国際原核生物命名規約に則って新属新種として提唱し、正式に認められました。このナノアーキアは好気条件で増殖できるため、ごく一般的な研究施設にて容易に培養可能であり、今後、モデル生物として広く利用されると期待できます。
本研究は、科学雑誌『International Journal of Systematic and Evolutionary Microbiology』オンライン版(8月22日付:日本時間8月22日)に掲載されました。

宿主アーキア(中央)の細胞表面に付着する寄生性ナノアーキア(左側の小さい細胞二つ)
背景
地球表層の環境は、1マイクロメートル(μm、1μmは100万分の1メートル)サイズの目に見えない小さなバクテリアおよびアーキアなどの微生物で満ちあふれています。その中でも、とりわけ小さな数百ナノメートルサイズの極小微生物が、全微生物種の15%以上を占めるともいわれています。しかし、それら極小微生物のほとんどがまだ実験室で培養できておらず、機能未知の「微生物ダークマター」の代表格として注目されています。
2002年に、世界で初めて極小微生物の一種であるナノアーキアが発見され、原核生物の分類上最も高次分類群である「門」のレベルで新規であることが明らかになり、世界中の研究者に衝撃を与えました。その後、極小微生物の培養がいくつか報告されましたが、一般利用可能なリソースとして公開されている極小微生物は、インドネシア産のたったの一種しかなく注1)、国産のものはありませんでした。微生物を培養してリソース化することは、科学の進歩を支える「再現性」の確保において極めて重要です。
そこで本研究では、極小微生物についてさらに理解を深めるために、国内に生息する極小微生物の培養およびリソース化を試みました。
注1)2022年1月17日プレスリリース「DPANN群に属する難培養性アーキアの培養に成功」
研究手法と成果
極小微生物は、土壌や地下水、温泉など地球上の幅広い環境に存在します。加藤真悟上級研究員らは、これまでに栃木県の酸性温泉を対象にして、微生物ハンティングを試みており、実際に新規性の高いアーキアの培養に成功しています注2)。同温泉の研究を通じて、この環境にナノアーキアが生息することは、培養に依存しない分子生物学的手法によって既に確認されていました。温泉水の化学分析および微生物群集解析の結果、この温泉に生息するナノアーキアは、酸素存在下でも増殖できる好気性の種であることが推定されました。そこで、この温泉水を植菌元として、好気条件下での培養実験を行いました。
培養を開始してから約1週間後、培養液を高倍率の光学顕微鏡で観察してみると、1μmサイズの一般的な微生物細胞の周りに、さらに小さな200~300nmサイズの小さな粒子が付着している様子が観察されました(図1)。遺伝子解析により、この培養液には、酸性温泉環境に一般的に生息する好熱好酸性アーキアのほかに、新しいナノアーキアが含まれていることが明らかになりました。この培養液をもとにして、さらに希釈・継代培養を繰り返すことで、1種類の好熱好酸性アーキアMJ1HA株と1種類のナノアーキアMJ1株のみを含む培養系の確立に成功しました。
ナノアーキアMJ1株およびMJ1HA株をそれぞれ異なる色で染色して、蛍光顕微鏡で観察すると、MJ1HA株の細胞の周りに付着している粒子が、ナノアーキアMJ1株であることが確認できました(図1)。MJ1HA株の細胞サイズが直径0.8~1.6μmであるのに対して、ナノアーキアMJ1株は直径200nm程度しかありませんでした。微生物を顕微鏡観察する際は一般的に200~400倍程度の倍率で観察しますが、それらの倍率ではナノアーキアMJ1株の存在すら認識できず、1000倍まで倍率を上げることで、ようやくその姿形を認識することができました。
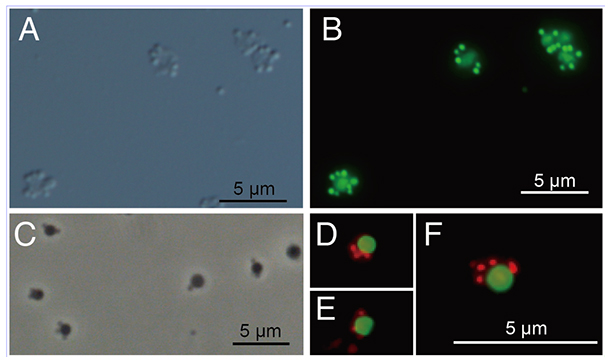
図1 ナノアーキアMJ1株と宿主アーキアMJ1HA株の顕微鏡観察像
(A)微分干渉顕微鏡観察像と(B)同視野の蛍光顕微鏡観察像。Bでは、どちらの細胞も緑色に見えている。大きめの細胞が宿主アーキアMJ1HA株で、その周りに付着している小さい細胞がナノアーキアMJ1株。(C)位相差顕微鏡観察像。(D-F)蛍光顕微鏡観察像。ナノアーキアMJ1株は赤色、宿主アーキアMJ1HA株は緑色に見えている。
注2)2019年6月11日プレスリリース「難培養アーキア(古細菌)の分離培養に成功」
さまざまな培養実験を実施しましたが、ナノアーキアMJ1株単独での増殖は確認できませんでした。また、MJ1HA株以外のアーキアとの共培養も試してみましたが、ナノアーキアMJ1株の増殖は確認できませんでした。そのため、ナノアーキアMJ1株が増殖するためには、宿主となるMJ1HA株の存在が必要不可欠であると考えられました。培養性状を詳しく調べたところ、ナノアーキアMJ1株および宿主MJ1HA株ともに、65~70℃、pH2.5付近を至適条件として増殖することが分かりました。
ナノアーキアMJ1株および宿主MJ1HA株の共培養系で、それぞれの株の増殖を継時的にモニタリングしたところ、培養初期では、ナノアーキアMJ1株の増殖により、宿主MJ1HA株の増殖が阻害されますが、その後は、お互いが緩やかに増殖していく様子が確認できました(図2)。一方で、宿主MJ1HA株は、ナノアーキアMJ1株が存在しなくても単独で増殖することができ、ナノアーキアMJ1株との共培養で見られた培養初期の増殖阻害は観察されませんでした。
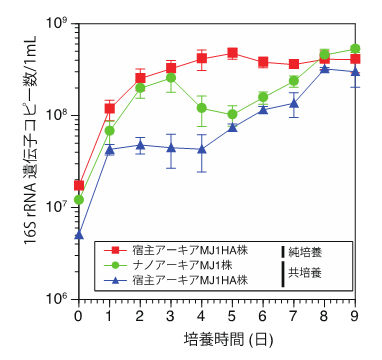
図2 ナノアーキアMJ1株と宿主アーキアMJ1HA株の増殖曲線
宿主アーキアMJ1HA株のみを含む純培養系(赤)と、ナノアーキアMJ1株と宿主アーキアMJ1HA株を含む共培養系(緑と青)において、それぞれの株の増殖を、16S rRNA遺伝子を標的にした定量PCR法により示した。
続いて、ナノアーキアMJ1株および宿主MJ1HA株の全ゲノム配列を決定しました。宿主MJ1HA株のゲノムサイズは2.28メガ塩基対(Mbp、1Mbpは100万塩基対)であり、単独で増殖するための遺伝子を全て保有していることが分かりました(図3)。その一方で、ナノアーキアMJ1株のゲノムサイズは0.67Mbpと極端に小さく、DNAやRNAのもととなる核酸や、細胞膜を構成する脂質、さらには全生物のエネルギー通貨であるATPなどの生命維持に必須の物質の生合成やエネルギー代謝に必要な遺伝子をほとんど保有していないことが明らかになりました(図3)。この結果は、先に記したナノアーキアMJ1株単独の培養ができなかったことと整合します。ナノアーキアMJ1株は、それら生命維持に必須の物質を、宿主アーキアMJ1HA株から奪い取って生育していることがゲノム解析からも支持されました。
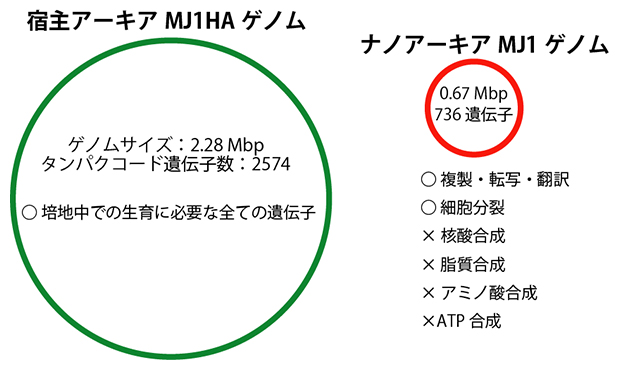
図3 ナノアーキアMJ1株と宿主アーキアMJ1HA株のゲノム
(左)宿主アーキアMJ1HA株のゲノムサイズとタンパクコード遺伝子の数。培養実験に使用した培地中で生育するための遺伝子を全て備えている。円の大きさは、ナノアーキアMJ1株のゲノムサイズと比較したときの相対的な大きさを示す。
(右)ナノアーキアMJ1株のゲノムサイズとタンパクコード遺伝子の数。DNAの複製・転写・翻訳や細胞分裂に関わる遺伝子はあるが、生命維持に必須の物質(核酸や脂質、アミノ酸、ATPなど)を合成するための遺伝子を持っていない。
それでは、ナノアーキアMJ1株はどうやってそれらの物質を宿主から奪っているのでしょうか。この疑問に答えるために、ナノアーキアMJ1株と宿主MJ1HA株が付着している状態の細胞を液体窒素で瞬間的に固めて、超薄切片を作製し、その切片を透過型電子顕微鏡で観察しました(図4)。その観察像を見る限り、ナノアーキアMJ1株が宿主MJ1HA株の細胞膜に穴を開け、そこから細胞質を吸い取っているように見えます。このような観察像はこれまでに報告例がなく、本研究によって、ナノアーキアの驚くべき生きざまを示唆する有力な証拠を提示することができました。
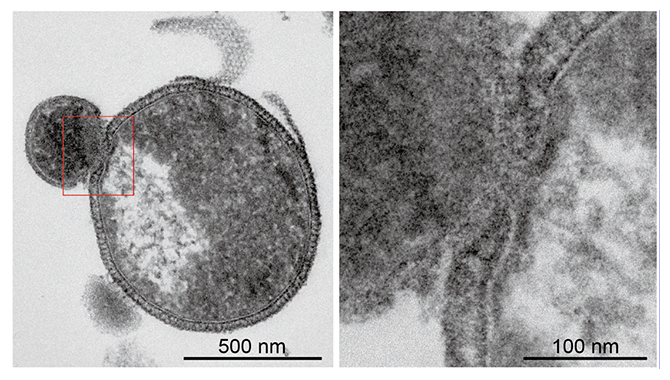
図4 超薄切片の電子顕微鏡観察像
(左)写真左上の小さいナノアーキアMJ1株が、宿主アーキアMJ1HA株に付着している様子。宿主アーキアの細胞質が吸い取られた結果、その部分の電子密度が薄くなって観察されていることが確認できる。
(右)左写真の赤枠の部分の拡大図。付着部分の細胞膜に穴が開いている様子が確認できる。
細胞膜を構成するコア脂質を調べたところ、ナノアーキアMJ1株から検出された成分は、宿主MJ1HA株から検出された成分と一致していましたが、その相対量に違いが確認できました(図5)。例えば、GDGT-5とGDGT-6というグリセロールテトラエーテルは宿主MJ1HA株からは多く検出されるものの、ナノアーキアMJ1株からはほとんど検出されてきませんでした。この結果は、ナノアーキアMJ1株は、膜を構成する脂質成分を宿主MJ1HA株から奪い取りますが、その中でもGDGT-3とGDGT-4を選択的に用いていることを示唆しています。
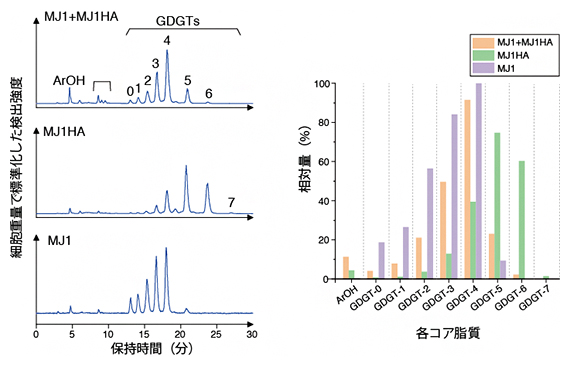
図5 ナノアーキアMJ1株および宿主アーキアMJ1HA株の細胞膜を構成するコア脂質成分
(左)抽出したコア脂質のクロマトグラム。上から、ナノアーキアMJ1株と宿主アーキアMJ1HA株を含む共培養系から細胞を集めた試料、宿主アーキアMJ1HA株の純培養系から細胞を集めた試料、共培養系からナノアーキアMJ1株の細胞のみを集めた試料の結果。
(右)左のクロマトグラムの結果に基づいて、各コア脂質の相対量を示したグラフ。ArOH:アーキオール。GDGT:グリセロールジアルキルグリセロールテトラエーテル。GDGTに続く数字は、イソプレノイド鎖に含まれる五員環の数を示す。
ゲノム配列に基づいて分子系統解析をしたところ、ナノアーキアMJ1株は、極小アーキアの中でも、2002年に海底熱水域から培養報告されたナノアーキアの一種Nanoarchaeum equitansや、陸上温泉から培養報告された種と比較的近縁であることが分かりました(図6)。これまでに報告されたナノアーキアは、全て嫌気性でしたので、ナノアーキアMJ1株は好気性のナノアーキアとして世界初の報告例です。その一方で、塩田から報告されたナノハロアーキアや、温泉や廃坑水から報告されたミクロカルドアーキアは、いずれも好気性かつ絶対寄生性もしくは共生性という点でナノアーキアMJ1株と類似しますが、それらとは門レベルで異なる系統群であることが確認されました(図6)。
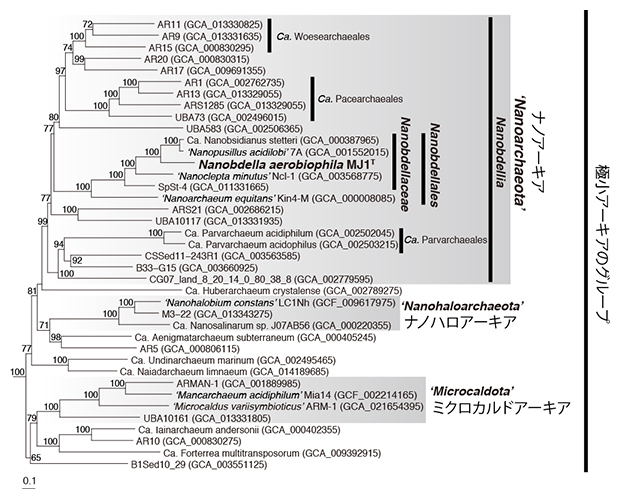
図6 ナノアーキアMJ1株の系統学的位置を示す系統樹
122個のマーカータンパク配列を結合したアライメントを基に、最尤法で作成した分子進化系統樹。日本語で示したグループ名は慣用名であることに留意されたい。
これまでに報告された極小微生物には、例えばNanoarchaeum equitansといった名前が暫定的に付けられていますが、正式に学名が認められている種は一つもありませんでした。そこで、共同研究グループは、ナノアーキアMJ1株を「Nanobdella aerobiophila」と命名し、国際原核生物命名規約に則って新属新種として提唱しました。さらに高次分類群として、新科Nanobdellaceae、新目Nanobdellales、新綱Nanobdelliaを提唱し、これらが正式に認められました。極小アーキア、さらには極小バクテリアも含めて、極小微生物の中で、正式に学名が認められたのは、本成果が世界初の例となります。ナノアーキアMJ1株(=JCM 33616T)および宿主アーキアMJ1HA株(=JCM 33617)は、各研究機関で利用可能な微生物リソースとして、理研の微生物材料開発室(JCM)で整備されています。
今後の期待
本研究では、好気条件で増殖できる絶対寄生性ナノアーキアの培養およびリソース化に世界で初めて成功しました。ナノアーキアMJ1株は、特別な設備や技術を必要とせず、一般的な研究施設において容易に培養ができることから、今後、同株を実験材料として、さまざまな新しい生命現象が実験的に明らかにされるものと期待できます。また、それらの実験から明らかにされるナノアーキアの生理性状を参考にして、さらなる極小アーキアの培養およびリソース化が促進されることでしょう。
絶対寄生性もしくは共生性の極小アーキアは、一般的な単細胞生物とウイルスの中間のような生命体として見なすことができ、また、分子系統樹上で最も根本付近から分岐したと推定されていることから、初期生命の姿を色濃く残しているとも考えられています。さらに、地球上に幅広く存在することが明らかにされていますが、そのほとんどがまだ培養されていない「微生物ダークマター」であり、その生態学的な役割はほとんど分かっていません。本成果を足がかりとして、今後極小アーキアの生理・生態の詳細が明らかになることで、大局的には、生命の初期進化や地球規模での物質循環の理解に貢献し、さらには「生命とは何か」という根源的な謎を紐解く手がかりを与えてくれると期待できます。
補足説明
1.アーキア(古細菌)
全生物は、真核生物(ヒト、動物、植物、カビなどを含む)、バクテリア(細菌)、そしてアーキア(古細菌)の三つに大きく分類されている。アーキアは、80℃以上の高温、20%以上の高塩分、pH1以下の強酸性などの極限環境を好む微生物を含む分類群である。その一方で、土壌や海水、海底堆積物といった非極限環境にも広く生息することが知られている。
共同研究グループ
理化学研究所 バイオリソース研究センター 微生物材料開発室
上級研究員 加藤 真悟(カトウ・シンゴ)
特別嘱託研究員 伊藤 隆(イトウ・タカシ)
テクニカルスタッフI 清水 美智留(シミズ・ミチル)
特別研究員(研究当時) 酒井 博之(サカイ・ヒロユキ)
開発研究員(研究当時) 雪 真弘(ユキ・マサシロ)
室長 大熊 盛也(オオクマ・モリヤ)
産業技術総合研究所 地圏資源環境研究部門 地圏微生物研究グループ
研究グループ付 金子 雅紀(カネコ・マサノリ)
東洋大学大学院 生命科学研究科 生命科学専攻
大学院生 小笠原 綾香(オガサワラ・アヤカ)
准教授 高品 知典(タカシナ・トモノリ)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業新学術領域研究(研究領域提案型)「超地球生命体を解き明かすポストコッホ生態学(領域代表者:高谷直樹)」、「ポストコッホ微生物資源の基盤整備(研究代表者:大熊盛也)」、理研横断プロジェクト「共生生物学プロジェクト(iSYM)」、発酵研究所研究助成金「ナノアーキアの検出と培養株確立の試み」(研究代表者:伊藤隆)および「我が国の温泉地における好熱性アーキアの地理分布、系統学的多様性および生理機能の解明」(研究代表者:加藤真悟)による支援を受けて行われました。
原論文情報
Shingo Kato, Ayaka Ogasawara, Takashi Itoh, Hiroyuki D. Sakai, Michiru Shimizu, Masahiro Yuki, Masanori Kaneko, Tomonori Takashina, Moriya Ohkuma, “Nanobdella aerobiophila gen. nov. sp. nov., a thermoacidophilic, obligate ectosymbiotic archaeon, and proposal of Nanobdellaceae fam. nov., Nanobdellales ord. nov., and Nanobdellia class. nov.”, International Journal of Systematic and Evolutionary Microbiology, 10.1099/ijsem.0.005489
発表者
理化学研究所
バイオリソース研究センター 微生物材料開発室
上級研究員 加藤 真悟(カトウ・シンゴ)
室長 大熊 盛也(オオクマ・モリヤ)
産業技術総合研究所 地圏資源環境研究部門 地圏微生物研究グループ
研究グループ付 金子 雅紀(カネコ・マサノリ)
報道担当
理化学研究所 広報室 報道担当
産業技術総合研究所 広報部 報道室

