
2023-12-11 理化学研究所,広島大学,大阪大学
理化学研究所(理研)生命機能科学研究センター 形態進化研究チームのサフィエ・エスラ・サルペル 学振特別研究員PD(日本学術振興会特別研究員-PD)、広島大学 大学院統合生命科学研究科 数理生命科学プログラムの藤本 仰一 教授(大阪大学 大学院理学研究科 生物科学専攻 招へい教授)、大阪大学 全学教育推進機構の北沢 美帆 講師(大阪大学 大学院理学研究科 生物科学専攻 兼任)らの共同研究グループは、放射相称動物[1]であるヒドロ虫[2]が体の周りに触手を配置する原理と、個体間に見られる放射相称性の多型を発見し、放射相称の触手配置および種内多型の仕組みを説明する数理モデルを提案しました。
本研究により、放射相称動物において器官が繰り返し配置されるルールや、異なる放射相称性が進化する仕組みについての理解が深まることが期待できます。
多くの生物は、その体に繰り返し構造を持ちます。ヒトを含む左右相称動物[1]では、脊椎など繰り返し構造を持つ器官の配置は2次元上での解釈および解析が可能です。しかし、体軸を中心に3次元的に器官が展開する放射相称動物の器官配置についての知見は限られていました。
今回、共同研究グループは、葉器官が3次元的に展開する植物体の解析手法をヒドロ虫の触手配置の分析に応用し、放射相称動物の器官が立体的に繰り返し配置される基本原理を発見しました。さらに、複数個体の触手の配置を定量的に解析した結果、個体のサイズに依存して異なるタイプの放射相称性が現れることが分かりました。また、数理モデルを用いたシミュレーションから、サイズに依存して放射相称のタイプが選択される仕組みの存在が示唆されました。
本研究は、科学雑誌『Frontiers in Cell and Developmental. Biology』(11月22日付)に掲載されました。

左右相称動物と放射相称動物、および植物に見られる繰り返し器官(オレンジ)
背景
動植物の形に見られる周期性(繰り返し構造)は、発生や成長の過程で広く現れる現象です。例えば、左右相称の脊椎動物では、周期的な器官である脊椎は体軸に沿って繰り返し配置され、これらの配置は前後および左右の2次元空間で解釈が可能であり、その形成メカニズムについて広く解析が行われてきました。一方、放射相称の動植物では、周期的な器官は体軸を中心に囲むように繰り返しながら配置されます。これらの中で一番解析されてきたのは、3次元空間で植物の茎を中心に配置された葉器官です。しかし、クラゲやヒドラなど放射相称の構造を持つ動物(刺胞動物[3])の周期的な器官配置に関する研究は限られており、どのようにしてこれらの動物が発生過程で放射相称性を確立するかは不明でした。
植物の葉器官の配置(葉序)は、器官間の距離と角度により、大きく二つのタイプに分類されます。一つは、植物の茎の同じレベル(茎の頂点から同じ距離)に複数の器官が付き、その繰り返しによって構成される、輪生と呼ばれるタイプ。もう一つは、器官が茎の長軸方向に一定の距離および角度の間隔を保ちながら繰り返し配置され、らせんパターンを形成するタイプです。またこれらのパターンには、種間の多様性に加えて、種内にも器官数と配置の多型(個体差)があることも報告されています。
刺胞動物のヒドロ虫は単純な円柱状の体を持ち、その周りに複数の触手器官を配置した放射相称性を確立しています。ヒドロ虫の放射相称性についてはこれまで、口の周りに少数(4~8本)の触手を持つヒドラ(Hydra vulgaris)などを対象として研究されてきました。しかし、多数(20~30本)の触手が体全体に複雑に配置された種について、触手がどのようなルールで配置されるのか、その配置が放射相称なのかはよく分かっていません。ヒドロ虫の一種であるCoryne uchidai[4](タマウミヒドラの仲間。以下、C. uchidai)は、植物の葉序のように、口(Oral)と口の反対側(Aboral)を結ぶ軸(O-A軸)を中心にした3次元空間で触手器官を繰り返し配置しています(図1上)。そこで、C. uchidaiを放射相称動物の新たなモデルとし、その触手配置を定量的に解析することで、動物においても植物と似たような器官配置の原則や多型が存在するかを調べました。
研究手法と成果
C. uchidaiは有性生殖と無性生殖を行い、無性生殖では親個体の口の反対側(反口側)が伸びて、そこから新たなポリプ[5]が出芽します。ポリプには複数の触手がリング状に形成され、3次元空間でO-A軸に沿って繰り返し配置されます。ライブイメージングで観察したところ、触手の形成はポリプの口の周りから始まり、ポリプの成長に伴って反口側に向かって順次配置されることが分かりました。また、一つのリングを構成する触手はほぼ同時に形成されていました(図1下)。

図1 C. uchidaiのポリプ形成のライブイメージング
(上)C. uchidaiの全体像。複数の触手が、ポリプの口側と口の反対側を結ぶ軸を中心に展開している。スケールバーは500マイクロメートル(μm、1μmは1,000分の1mm)。
(下)出芽したポリプの成長を捉えたライブイメージング。タイムスケールは時:分を示す。スケールバーは250μm。
次に、触手の空間配置を定量的に分析するため、植物の形を記述する測定法を応用し、口を茎の頂点、触手を葉序と見なしてそれらの配置を測定しました。口からの距離が近い順で触手を並べ替え、それぞれの触手間の距離を分析しました。その結果、口の周りに4本の触手で形成される最初のリング(以下、第1リング)から始まって、触手が4本ごとに大きく間隔を開けて配置されていることが明らかとなりました。これは、植物の葉の輪生配置と似た特徴です(図2左)。輪生配置では、輪生内の葉は規則的(等角度)に並びます。そこで、C. uchidaiの第1リングの触手間の角度を計測した結果、平均角度は92.84°と直角に近く、四放射状の対称性(四放射相称)を示すことが分かりました(図2右)。
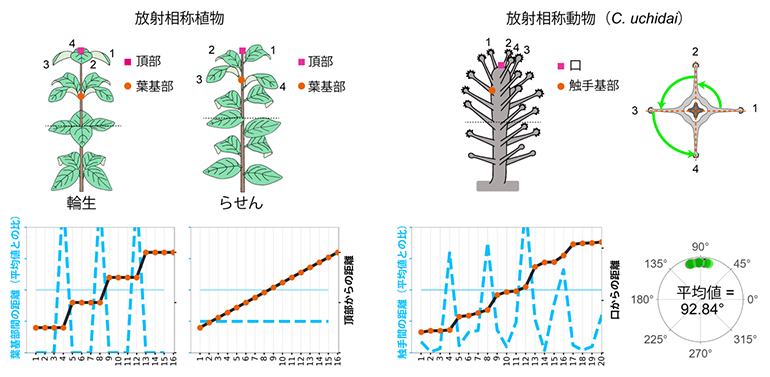
図2 放射相称植物の輪生配置と放射相称動物の触手配置の共通性
(左)植物の模式図(上)と、葉の配置を測定したグラフ(下)。植物の葉器官の配置(葉序)は、葉が植物の茎の周りに同じレベル(頂点から同じ距離)で繰り返し配置される輪生と、葉が縦方向に一定の距離および角度の間隔を保ちながら繰り返し配置されるらせんの2パターンに大別される。頂点から順番に葉に番号を振り、葉と葉の間の距離をそれぞれの葉の基部(オレンジの点)間の距離として計測し(青の破線)、また、頂点からそれぞれの葉の基部までの距離を黒の実線でつないだ。輪生では、4枚の葉ごとにまとまったリングを形成していることがグラフ上で示された。
(右)C. uchidaiの触手配置を、植物と同様の手法で解析した結果。模式図下のグラフにおいて、青の破線は触手(の基部)間の距離、黒の実線は口からの触手(の基部)までの距離を示す。グラフのパターンは輪生と類似し、4本の触手がまとまってリングを形成していることが示された。右端は触手を上から見た模式図で、触手間の角度の平均値は92.84°とほぼ直角となり、四放射状の対称性(四放射相称)であることが示された。
多くのポリプを調べた結果、ほとんどのポリプは第1リングに4本の触手を持っていましたが(41例)、3本(13例)と5本(12例)の触手を持っているものも発見しました。第1リングに3本の触手を持つ個体では、隣同士の触手間の平均角度は約120°であり、三放射相称を示していました。同様に5本の触手を持つリングでは、触手間の平均角度は約72°の五放射相称を示していました(図3)。さらに、上下に連続するリング間の関係に着目すると、三放射状、四放射状、五放射状のポリプにおいて上下のリングの触手間の角度はリング内の触手間の角度の半分で、すなわちそれぞれ約60°、45°、36°でした。これは、上下のリングは交互(互い違い)になるように触手を配置していることを示し、輪生植物の葉序に共通する特徴です。以上の結果は、C. uchidaiでは、輪生葉序に似て同一リング内の触手は等角度で、連続するリング間で触手を交互に配置させる「放射状の対称性」が全身にわたって存在することを示しています。
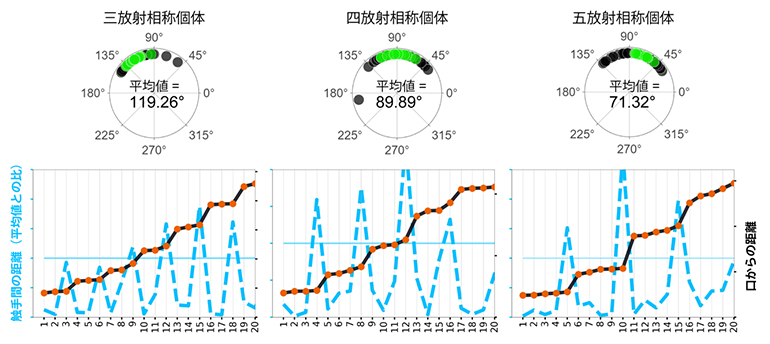
図3 C. uchidaiに見られる放射相称性の個体差
(上)第1リングの触手の数が3本(左)、4本(中)、5本(右)のポリプそれぞれについて、触手間の角度を測定した結果を円上の角度で示した。それぞれの平均値は約120°、90°、72°となり、三放射相称、四放射相称、五放射相称の対称性を持つことが分かった。
(下)それぞれの放射相称性を持つ触手の配置を示したグラフ。青の破線は触手間の距離、黒の実線は口からの触手までの距離を示す。三放射相称個体は3本の触手がまとまってリングを形成し、五放射相称個体は5本の触手がまとまってリングを形成していることを示す。
上記のように、C. uchidaiの触手配置には種内変異が見られ、複数の放射相称のタイプが存在します。ではこれらの違いは、どのように選択・決定されるのでしょうか。C. uchidaiと同じ刺胞動物の一種であるヒドラ(Hydra vulgaris)は、やはり個体によって異なる数の触手が口の周りにリング状に配置されることが知られています。ヒドラでは、触手の数はポリプのサイズと正の相関を示し、サイズが大きいポリプほど多くの触手を持っていることが報告されています注1)。しかし、器官の数の変異と関連する「対称的な配置の多型」がサイズに依存するかどうかは、明らかにされていません。そこで、C. uchidaiにおける個体サイズと対称性タイプの関係を調べたところ、ポリプの直径が大きいほど相称面の数が増える(三放射→四放射→五放射)ことが分かりました。これは、個体サイズが放射相称のタイプとも相関しており、対称性の多型がポリプの直径のサイズの違いから生じている可能性を示しています。
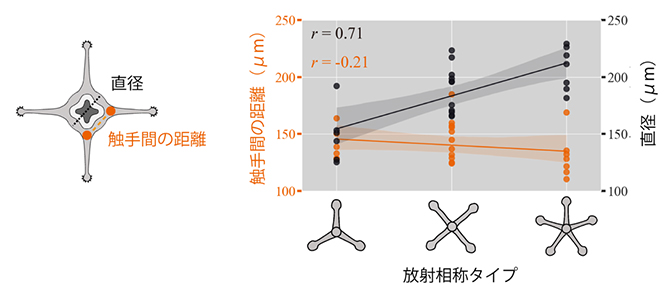
図4 C. uchidaiに見られる放射相称タイプと個体サイズ
(左)ポリプの第1リングの直径と触手間の距離を示す模式図(断面図)。
(右)放射相称のタイプ(横軸)と、触手間の距離(左の縦軸)、ポリプの直径(右の縦軸)の関係を示したグラフ。相称面の数の増加(三放射→四放射→五放射)はポリプの直径と正に相関するが、触手間の距離とは相関しない。
最後に、周期的な触手配置と、サイズの多様性と相関した相称性の多型が出現する機構を推測するために、上記の観察事実を説明可能な数理モデルを構築しました。ヒドラについて行われた先行研究では、触手の発生に関与する三つの拡散性因子が仮定され、それぞれ活性化因子A、抑制因子B、および側方抑制[6]因子Cとして機能します。活性化因子Aと抑制因子Bはどちらもヒドラの口領域から分泌されます。一方の側方抑制因子Cは、各触手から分泌されます。これらの因子はいずれも分泌場所から拡散して濃度勾配を形成し、触手の形成を促進あるいは抑制します。このモデルでは、ある領域が以下の条件を満たす場合、触手になると設定します。
①活性化因子Aの濃度(a)が閾値(Ta)を超えている(a > Ta)
②抑制因子Bの濃度(b)が閾値(Ti)を下回っている(b < Ti)
③側方抑制因子Cの濃度(c)が閾値(Tti)を下回っている(c < Tti)
①と②の条件により口から一定の距離を保った触手配置が行われ、③の条件により触手がお互いの配置を抑制し、等角度に触手が配置されます。
これらヒドラの先行研究で報告されたモデル注2)の特徴、すなわち口から一定の距離の領域で等角度に並ぶ触手配置はC. uchidaiと共通しています。しかしC. uchidaiは口の周りの第1リングに加えて、体全体に複数のリングを配置しています。そこで共同研究グループはさらに、
④活性化因子Aの最大値で触手が配置される
という条件を加えたモデルへと拡張し、C. uchidaiを模した円筒形のポリプ体の単一細胞層表面でこの数理モデルをシミュレーションしました(図5上)。すると、触手は一定の距離を保ちつつ口の近くに第1リングを作り始めました。これは、活性化因子Aの濃度が閾値(Ta)を超え、かつ抑制因子Bが閾値(Ti)を下回っていることに起因しています。また、第1リングに4本の触手が順次できる際、隣同士の触手間の角度が約90°(90.69°)になりました。これは、抑制因子Cに起因しています。活性化因子Aは体全体で機能し、各触手からは側方抑制因子Cが拡散します。このため、4本の触手ごとの周期で複数のリングが形成され、上下に連続したリングで交互に触手が配置されました(図5下中)。さらに、ポリプ体の直径が増加するにつれて五放射相称の個体が現れ(図5下右)、直径が減少すると三放射相称の個体が現れることも再現できました(図5下左)。
以上の結果から、触手の配置を制御する活性化因子と抑制因子の組み合わせにより、等角度の触手で構成されたリングが周期的に複数現れる放射相称性が出現し、個体のサイズと相関する対称性の多型を再現できることが分かりました。本研究で提示した数理モデルは、3次元空間での器官配置に基づく対称性が、個体のサイズに応じて制御されることを示唆しています。
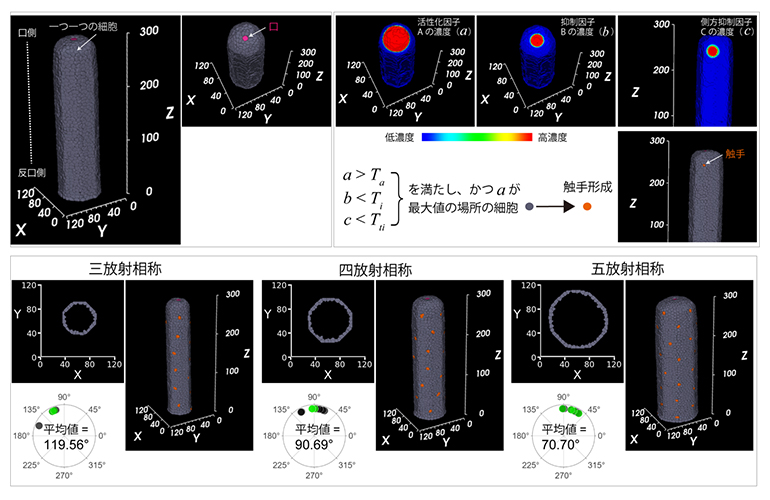
図5 C. uchidaiの触手配置数理モデルのシミュレーション
(上)C. uchidaiを模した円筒形モデル(左)。ポリプ体の単一細胞層を灰色のドットで示す。シミュレーションの初期値として、円柱の頂点に口(ピンク)を置いた。この時、口や触手から分泌される因子の濃度勾配をa(活性化因子A)、b(抑制因子B)、c(側方抑制因子C)の図の擬似カラーで示す(右)。シミュレーションの結果、触手(オレンジ)ができた。
(下)ポリプの直径を変えた時のシミュレーション結果。直径の大きさに依存して相称面の数が増加(三放射→四放射→五放射)する現象が再現できた。
注1)Parke, H. H. (1900). Variation and regulation of abnormalities in Hydra. Arch. für Entwicklungsmechanik Org. 10, 692-710.
注2)Meinhardt, H. (2012). Modeling pattern formation in hydra: a route to understanding essential steps in development. Int. J. Dev. Biol. 56, 447-462.
今後の期待
放射相称動物は、その進化的な起源や体の成り立ちに関して多くの謎が残されています。本研究は、刺胞動物のC. uchidaiをモデルとし、複数の触手が等角度に配置されたリングを作ることと、このリングの周期的な繰り返しが放射相称の触手配置の基本原理であることを明らかにしました。さらに、対称性のタイプにおける多型(三放射相称、四放射相称、五放射相称)はポリプの直径と相関することも見いだしました。本研究で確立した定量的な解析方法および触手配置の数理モデルは他の放射相称動物にも適用可能であり、今後、放射相称性の多様性や進化的移行と個体サイズの関係についての新たな知見が得られると期待できます。
補足説明
1.放射相称動物、左右相称動物
動物の体の体制について、前と後、背と腹が区別され、外見的に左側と右側の形が等しい体を持つものを左右相称動物と呼ぶ。一方、一つの対称軸を含む相称面が複数存在するものを放射相称動物と呼ぶ。
2.ヒドロ虫
刺胞動物門の中のヒドロ虫綱に属する種の総称。無性生殖により多数の個体がつながった群体を形成する。
3.刺胞動物
動物分類の最も大きな単位「門」のうち、クラゲ、ヒドラ、サンゴ、イソギンチャクなどを含むグループ。
4.Coryne uchidai
世界各地に分布するタマウミヒドラ(Coryne pusilla)の仲間。本研究で用いたCoryne uchidaiは、兵庫県明石市で採取したもの。
5.ポリプ
刺胞動物門の動物で現れるライフサイクルステージの一つ。固着生活に適した形態をとる。
6.側方抑制
多細胞生物の発生過程では、ある細胞が分泌した因子の拡散により、他の細胞の分化が阻害される例がよく知られており、この現象を側方抑制と呼ぶ。
共同研究グループ
理化学研究所 生命機能科学研究センター 形態進化研究チーム
学振特別研究員PD サフィエ・エスラ・サルペル(Safiye Esra Sarper)
(日本学術振興会 特別研究員-PD)
テクニカルスタッフⅠ 仲西 珠美(ナカニシ・タマミ)
広島大学 大学院統合生命科学研究科 数理生命科学プログラム
教授 藤本 仰一(フジモト・コウイチ)
(大阪大学 大学院理学研究科 生物科学専攻 招へい教授)
大阪大学 全学教育推進機構
講師 北沢 美帆(キタザワ・ミホ)
(大阪大学 大学院理学研究科 生物科学専攻 兼任)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業特別研究員奨励費「タテジマイソギンチャクの胃の分節構造に現れる対称性の種内多型(研究代表者:サフィエ・エスラ・サルペル)」、同挑戦的研究(萌芽)「刺胞動物の放射対称性と左右対称性を調節する原理の構成的理解:実験と数理モデル(研究代表者:藤本仰一)」、同新学術領域研究(研究領域提案型)「器官配置の周期を構成し変調するメリステム動態の理論生物学(研究代表者:藤本仰一)」による助成を受けて行われました。
原論文情報
Safiye E. Sarper, Miho S. Kitazawa, Tamami Nakanishi, and Koichi Fujimoto, “Size-Correlated Polymorphisms in Phyllotaxis-Like Periodic and Symmetric Tentacle Arrangements in Hydrozoan Coryne uchidai”, Frontiers in Cell and Developmental. Biology, 10.3389/fcell.2023.1284904
発表者
理化学研究所
生命機能科学研究センター 形態進化研究チーム
学振特別研究員PD サフィエ・エスラ・サルペル(Safiye Esra Sarper)
(日本学術振興会 特別研究員-PD)
広島大学 大学院統合生命科学研究科 数理生命科学プログラム
教授 藤本 仰一(フジモト・コウイチ)
(大阪大学 大学院理学研究科 生物科学専攻 招へい教授)
大阪大学 全学教育推進機構
講師 北沢 美帆(キタザワ・ミホ)
(大阪大学 大学院理学研究科 生物科学専攻 兼任)
報道担当
理化学研究所 広報室 報道担当
広島大学 広報室
大阪大学全学教育推進機構等事務部総務係
")
