
2018/10/01 自然科学研究機構 基礎生物学研究所,名古屋大学
被子植物の受精の過程では、花粉から花粉管が長く伸長し、卵細胞のもとへ精細胞が運ばれることが必須です。花粉管が正常に伸長するためには、ANXURに代表されるいくつかの受容体タンパク質が花粉管の先端部に局在してはたらくことが必要ですが、その局在化のしくみはこれまで分かっていませんでした。
今回、基礎生物学研究所の室啓太特別協力研究員および上田貴志教授らの研究グループは、理化学研究所の中野明彦光量子工学研究センター副センター長、名古屋大学の東山哲也教授のグループと協働し、花粉管の伸長に必要なタンパク質ANXURを花粉管先端へと正しく局在させるために必要な分子を発見しました。この分子(PICALM5aおよびPICALM5b)は、膜に埋め込まれたタンパク質の輸送に関わるANTHタンパク質の一種で、花粉管の先端よりやや基部の亜頂端領域で形成される輸送小胞にANXURタンパク質を積み込むはたらきを担っていました。PICALM5aとPICALM5bの機能を失わせると、ANXURは花粉管の先端に局在することが出来なくなり、花粉管は伸長途中で破裂してしまいました。
一方、花粉管を正しい方向に導くための別の受容体タンパク質の輸送小胞への取り込みにはPICALM5aとPICALM5bは必要がないことも示されました。ANTHドメインを持つタンパク質は陸上植物の進化の過程で劇的に数が増えていますが、その生物学的な意味はこれまで分かっていませんでした。本研究により、ANTHタンパク質の機能の多様化が、植物の生殖の進化と深く関連していることが示されました。
本研究成果は2018年9月26日付でCommunications Biology誌に掲載されました。

図:野生型とpicalm5a picalm5b 二重変異体の花粉管の先端。
二重変異体の花粉管は伸長途中で破裂してしまう。
【研究の背景】
被子植物は花粉を介した有性生殖を行います。めしべの柱頭に受粉した花粉は、柱頭から水や養分を吸収し、花粉管という細長い管状の構造を発芽・伸長させます。花粉管は花柱を通過し、伝達組織内を伸長し、胚珠からの誘導を受けて胚珠へ到達します。花粉管の中には2つの精細胞が含まれており、花粉管の中を胚珠まで移動し、ひとつは卵細胞と、もうひとつは中央細胞と融合します。このプロセスは植物の生殖に留まらず、穀物となる種子の形成や異種間のかけ合わせによる作物の育種など、ヒトの生活にも深く関わる重要な現象です。
先行研究により、花粉管の伸長はさまざまな花粉管特異的な膜タンパク質によって制御されていることが明らかになっていました。たとえば、シロイヌナズナのPRK6は、胚珠から分泌されるLUREペプチドに結合することで、花粉管の胚珠への誘導において機能することが報告されています。また、ANXURは、花粉管自身が分泌するペプチドであるRALF4およびRALF19と相互作用して花粉管の破裂を防ぐ一方で、胚珠でつくられるRALF34と相互作用すると、花粉管の破裂を引き起こすことが分かっています。この仕組みは、適切なタイミングで花粉管を破裂させることによって、受精に必要な精細胞の放出を制御していると考えられます。
これらの膜タンパク質はいずれも花粉管の先端付近に局在していますが、高速で伸長する花粉管において、どのようにその局在を維持しているかについてはよく分かっていませんでした。花粉管の先端の細胞質には分泌小胞が蓄積しており、伸長のために必要な膜や細胞壁の成分をエキソサイトーシスにより供給しています。また、先端から少し離れた亜頂端領域の細胞膜からは、クラスリン依存性エンドサイトーシスにより物質の取り込みが盛んに行われており、その多くは、すばやく先端に送り返されるということが知られています。ANTHタンパク質は、クラスリン被覆小胞の形成の際に、細胞膜や積み荷タンパク質とクラスリンとの結合を仲介するアダプターとしてはたらいていると考えられている分子です。ANTHタンパク質は、シロイヌナズナゲノム中に多数コードされており、その機能はメンバー間で多様化していると考えられていました。しかし、植物のANTHタンパク質の生理機能については、これまでほとんど明らかにされていませんでした。また、ANTHタンパク質の多くが花粉で発現していることから、花粉や花粉管で特に重要な役割を担うと推測されていましたが、個々のメンバーがどのような積み荷タンパク質のクラスリン小胞への積み込みに関わるのかについては、全く分かっていませんでした。
【研究の成果】
この研究の最大の成果は、植物でほとんど明らかになっていなかったANTHタンパク質の生理的機能を、世界に先駆けて明らかにしたことです。
本研究ではまず、花粉で発現するANTHタンパク質PICALM5aとPICALM5bの二重変異体において、それぞれの鞘に形成される種の数が野生型と比較し、著しく減少していることを見いだしました(図1)。詳しく調べると、picalm5二重変異体の花粉管は伸長中に破裂してしまうため(図2)、種子の数が減少していることがわかりました。蛍光タンパク質であるGFPを融合させたPICALM5aやPICALM5bは、花粉管の先端よりやや基部側(亜頂端領域)の細胞膜に局在し、クラスリン軽鎖と非常に良く共局在したことから、PICALM5は花粉管の亜頂端領域からのクラスリン依存性エンドサイトーシスではたらいていることがわかりました。さらに、野生型では花粉管の先端に集中して局在するANXURが、picalm5二重変異体の花粉管では広範囲の細胞膜上に拡散してしまうことも見いだしました(図3)。これらの結果から、PICALM5はANXURをクラスリン被覆小胞に積み込み、細胞膜から取り込んで先端に送り返すリサイクリング経路に乗せる役割を担っていると考えられます。
一方、花粉管の誘因を担うPRK6については、picalm5二重変異体においても野生型と同様に花粉管の先端の細胞膜に局在していました。さらに、二重変異体の花粉管もLUREに正常に誘引されることも分かりました。これらの結果から、花粉管の伸長ではたらく二種類の膜タンパク質は、それぞれが異なるアダプタータンパク質により、リサイクリングのための小胞に積み込まれ、花粉管の先端へ局在化していることが明らかとなりました。

図1 picalm5a picalm5b 二重変異体では鞘に含まれる種子の数が
野生型と比較し少ない。
図2:野生型とpicalm5a picalm5b 二重変異体の花粉管の先端。
二重変異体の花粉管は伸長途中で破裂してしまう。
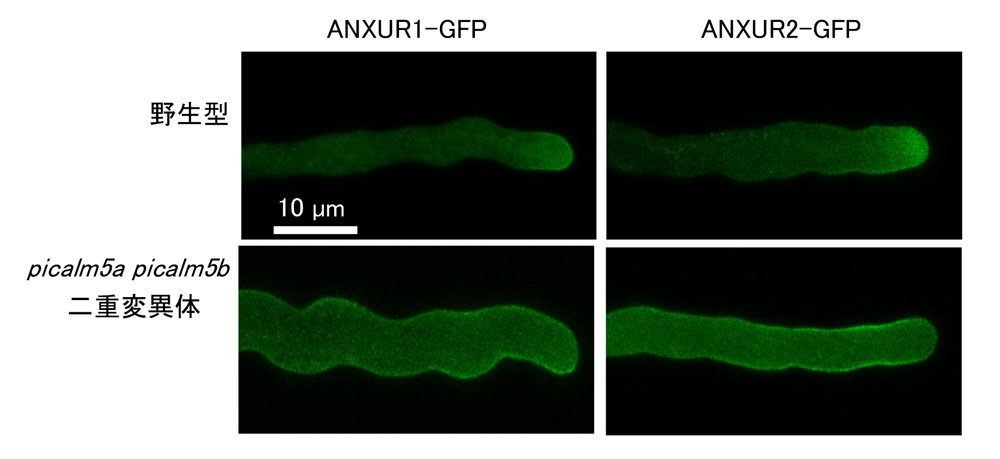
図3:GFPを融合したANXURタンパク質の花粉管での局在。
野生型の花粉管では先端付近にANXURが集まっているのに対し、
picalm5a picalm5b二重変異体では広範囲の細胞膜に拡散している。
【今後の展望】
今回の研究から、ANTHタンパク質の一種であるPICALM5aとPICALM5bが、花粉管の正常な伸長に必要なANXURタンパク質の花粉管先端部への局在に関わることが明らかになりました。シロイヌナズナには花粉での発現が報告されているANTHタンパク質が他にも多く存在します。今後、花粉管ではたらく他のANTHタンパク質の機能を明らかにすることで、ANTHタンパク質の多様化と植物の生殖システムの進化の関係についてさらなる知見が得られるものと期待されます。
【発表雑誌】
雑誌名:Communications Biology 2018年9月26日 掲載
論文タイトル:ANTH domain-containing proteins are required for the pollen tube plasma membrane integrity via recycling ANXUR kinases
著者:Keita Muro, Kumi Matsuura-Tokita, Ryoko Tsukamoto, Masahiro M. Kanaoka, Kazuo Ebine, Tetsuya Higashiyama, Akihiko Nakano, and Takashi Ueda
DOI:10.1038/s42003-018-0158-8
【研究グループ】
本研究は、基礎生物学研究所 細胞動態研究部門の室啓太特別協力研究員と上田貴志教授らのグループを中心として、理化学研究所の中野明彦光量子工学研究センター副センター長,名古屋大学の東山哲也教授らのグループからなる共同研究チームにより実施されました。
【研究サポート】
本研究は、科学研究費助成事業(25221103,16H06464,15H01231,17H05850,24114003,15H04382,17K19412,18H02470),三菱財団,および山田科学振興財団などの支援を受けて行われました。
【本研究に関するお問い合わせ先】
基礎生物学研究所 細胞動態研究部門
教授 上田 貴志 (うえだ たかし)
名古屋大学トランスフォーマティブ生命分子研究所
教授 東山 哲也(ひがしやま てつや)
【報道担当】
基礎生物学研究所 広報室
名古屋大学総務部総務課広報室

