2022-12-23 東京大学
小原 晶奈(神奈川大学 理学部4年)
小川 麻里(安田女子大学 准教授)
尾山 洋一(釧路市教育委員会 マリモ研究室次長)
鈴木 祥弘(神奈川大学 准教授)
河野 優(生物科学専攻 特任助教)
発表のポイント
- 結氷期間の短縮(消失)がもたらす冬の厳しい環境は、マリモ藻類に深刻なダメージを与える可能性がある。
- マリモ藻類には光合成系に未知の低温光修復機構があることや、修復の限界を明らかにした。
- 特別天然記念物「阿寒湖のマリモ」の生態解明に迫る新たな視点を与え、保護・保全の指針となる。
発表概要
北海道阿寒湖には、緑藻類Aegagropila linnaei(マリモ)の糸状体が集まって作る球状集合体が群生している。夏から冬にかけて大きな水温差を経験するマリモの生育様式は未だ不明な点が多い。水温が1-4 ℃にまで低下する冬期、太陽の強い日差しは湖面の凍結と積雪によって遮られるため、マリモはほとんど光が届かない暗い環境下で生育している。ところが、地球規模の温暖化が進み、湖面が結氷しなくなると、マリモは危険な活性酸素生成につながる低温・強光環境に曝される可能性が高い。東京大学大学院理学系研究科の河野優特任助教らは、結氷消失によって予想される低温・強光が阿寒湖のマリモに与える影響を解析した。マリモの糸状体細胞は、これまでに知られていない光修復機構を持っており、強光で生じた損傷を低温下で速やかに修復できること、結氷消失後の生息地に予想される自然環境下では十分な修復が難しく、細胞が枯死することを明らかにした。冬期の結氷消失はマリモに重大な損傷を与える可能性、およびマリモの生存における結氷の重要性が示唆された。温暖化の影響について、具体的に警鐘をならす結果となった。
発表内容
北海道東部に位置する阿寒湖では、緑藻類Aegagropila linnaei(マリモ)の糸状体が集まって球状の集合体を形成することが知られている(図1a)。以前は世界中の多くの湖沼でマリモの生育が確認されていたが、1900年代以降の生育環境の変化により、その多くが消失もしくは減少してしまった。中でも、最大直径30 cmに及ぶ巨大なマリモが観測されるのは世界でも阿寒湖だけであり、その希少性と保護の観点から、国の特別天然記念物に指定されている。阿寒湖では、夏期の水温は20-25℃ となり、晴天時、水中には太陽からの比較的強い直達光が差し込む。一方、冬期の水温は1-4℃にまで低下し、結氷と積雪(図1b)により太陽からの直達光は水中にほとんど届かない。夏と冬で全く異なる阿寒湖の温度・光環境への対応は、マリモの生存に不可欠である。近年、地球規模の環境変化の影響で、阿寒湖の結氷期間が短縮傾向にあり、マリモへの影響が懸念されている。今後、温暖化により冬期の結氷が失われた場合、水中に直接入射する太陽光により光強度(注1)は上昇する一方、結氷による断熱効果が失われた湖水の温度は比較的低温のまま維持されると予想される。低温下での強光曝露は、活性酸素(注2)の生成を促し、葉緑体光合成膜の光合成電子伝達成分(注3)を阻害する(図2)。しかし、マリモの光合成に着目した研究は少なく、マリモの年間を通した基本的な光合成の動態すら不明な点は多い。東京大学大学院理学系研究科の河野優特任助教らは、温暖化による結氷消失はマリモの光合成系に甚大な阻害を与えると考え、低温下の光合成の実態解明に取り組んだ。

図1:(a) 阿寒湖に生育する球状集合体マリモ。設置してある装置は光の強さを測るための光量子計。(b) 1-3月の阿寒湖の様子。湖面は氷と積雪により覆われる。

図2:上段:結氷と積雪により強い太陽光から保護されている場合(左図)と、結氷消失し、低温・強光に曝された場合(右図)のイメージ。
下段:低温下、弱光下での光合成(左図)と強光下での光合成(右図)。光合成は、葉緑体の光合成膜にある電子伝達成分のひとつである光化学系II (PSII)が光励起され、もうひとつの光化学系I (PSI)に電子が流れることで進行する。光が強すぎると、光化学系IIは壊れてしまう(右図の「損傷」)。通常、藻類や植物は壊れた光化学系IIを修復する機構を持っており(右図の「修復」)、その過程の一部では酵素が関与する。そのため、酵素活性が低くなる低温下では修復もほとんど起きない、もしくは非常にゆっくりとしか起きないとされている。しかし、マリモは低温下でも高い光修復能を示した(右図)。
阿寒湖が結氷している3月のある晴れた日に、氷に約3 m四方の穴を開けて、マリモ群落直上の水温と光強度を測定した。この結果をもとに、文化庁の許可を得て採集したマリモ球状体を用いて検証実験を行った。マリモ球状体の表面から細胞を傷付けないように丁寧に糸状体を引き抜き、この糸状体を、2℃下で強光にさらした。糸状体細胞の光合成は短い強光照射で容易に阻害(強光阻害;(注4))されたが、この後に比較的弱い光を当てることで阻害前のレベルまで速やかに回復した(図3a, 光修復; (注5))。これまで、損傷した光合成装置の修復は低温下では起きにくいとされていたことから、マリモ細胞には未知の修復機構が存在することが示された。一方、マリモ糸状体を結氷消失後の生息地で予想される疑似自然光環境下に置いたところ、光合成の阻害からの回復は見られなくなった(図3b)。さらに、数日間観測を続けたところ、糸状体細胞は枯死してしまった(図3c, d)。
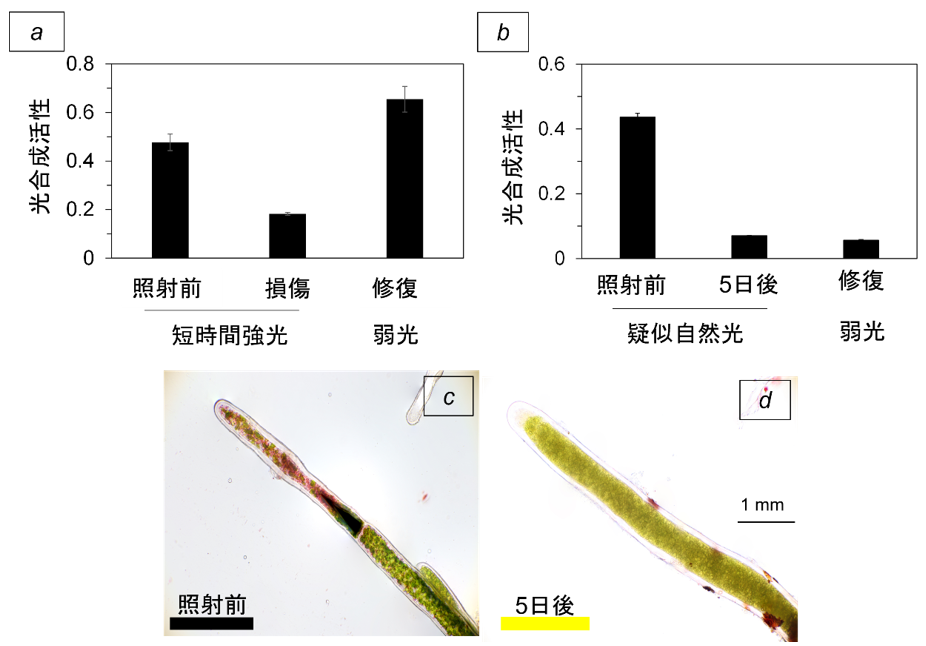
図3: ( a ) マリモの糸状体細胞(図中の)「照射前」に、結氷消失後に想定される強さの強光を数時間当てたとき「損傷」、および損傷細胞に弱い光を当てた「修復」の光合成活性の比較。
( b ) 結氷消失後の生息地で予想される疑似自然光環境にさらしたときの光合成活性の比較。光が点灯している明期(12時間)の光の強さは、暗闇(日の出,「照射前」)から徐々に光強度が上がっていき、光点灯6時間後に( a )と同じ光強度(南中)に達した後、6時間かけてまた暗闇(日の入)まで低下するように設定した。夜に相当する暗期も12時間に設定した。これを5日間繰り返した「5日後」と、その後に光強度を半分にした同周期の光を1週間当て続けた「修復」の光合成活性を比較した。
(c と d) ニュートラル・レッド染色法によって細胞の生死を確認した。生細胞は赤く染まるが( c ) 、死細胞は染まらず、緑のままである ( d )。 ( a )~( d )の実験はすべて、2℃下で行った。
マリモの糸状体細胞は、低温・強光の環境に一定時間は耐えられるが、自然環境下で予想される長時間の低温・強光には耐えられないことが明らかになった。結氷がマリモの生存に重要であることを示唆した本研究は、湖沼の生物への温暖化の影響に警鐘を鳴らすものである。また、国の特別天然記念物「阿寒湖のマリモ」(注6)保護の具体的な対応策提案にもつながる。今回の知見はマリモ糸状体細胞を用いて得られたものであり、球状集合体としての応答も詳細に調べる必要がある。今後は、阿寒湖の水温/光環境の経時観測を行うと同時に、球状体集合体としてのマリモの光合成と損傷の実態解明に取り組む。また、光修復の低温適応機構の解明にも取り組む。
本研究は、科研費「若手研究(課題番号:21K15118)の支援により実施された。
発表雑誌
- 雑誌名
International Journal of Molecular Sciences論文タイトル
Effects of high irradiance and low water-temperature on photoinhibition and repair of photosystems in Marimo (Aegagropila linnaei) in Lake Akan, Japan著者
Akina Obara, Mari Ogawa, Yoichi Oyama*, Yoshihiro Suzuki*, Masaru Kono,*
*責任著者DOI番号
10.3390/ijms24010060
用語解説
注1 光強度
光合成反応では、光を粒子(光量子)としてみたときに、光量子がどの程度のエネルギーを持っているのかではなく、光量子の数が重要となる。そのため、光合成分野では、波長400-700 nmの光量子数を基本とした単位、光量子束密度(μmol photons m-2 s-1)を使う。一般的に、室内の蛍光灯下が10 μmol photons m-2 s-1程度、真夏の直射日光がおよそ2000 μmol photons m-2 s-1程度になる。冬期、晴れた日の阿寒湖面で1500 μmol photons m-2 s-1程度、結氷後の湖底で10 – 40 μmol photons m-2 s-1程度、氷を除去して強い太陽光が届くようになった場合の湖底で500 – 800 μmol photons m-2 s-1程度であった。
注2 活性酸素
光合成に利用できない行き場を失なった過剰な光エネルギーは、葉緑体の中で活性酸素を作り出す。活性酸素は、通常の酸素とひとつの電子が反応してできる。活性酸素は反応性が高く、光合成装置を破壊してしまう。
注3 光合成電子伝達
葉緑体の光合成膜で起こる光合成電子伝達反応は、2つの光化学系IIとI(それぞれ、PSIIとPSI)の励起を駆動力として起こる(図2下段も参照)。光化学系IIを電子伝達の出発点とし、光化学系Iに電子が渡り、最終的に還元力が生成される。光合成生物はこの電子の流れを利用して、水素イオンを光合成膜の内側に取り込む。電子の流れと水素イオンの流れによってできた光合成膜を介したエネルギー差を利用してATPが合成される。光合成電子伝達反応により得られた還元力とATPを利用して、二酸化炭素を固定する。↑
注4 強光阻害
強光による光合成機能が低下する現象のこと。光合成速度は照射される光の強さに比例して高くなる。しかし、ある程度の強度に達すると光合成速度は飽和する。光合成を飽和させるレベルよりも強い光が照射されると、過剰な光エネルギーが発生する。過剰な光エネルギー下では反応性の高い活性酸素種が生成され、これらが光合成装置を損傷する。強光阻害は光合成生物の生育を妨げる大きな要因となる。 ↑
注5 光修復
強光阻害による光合成の低下は、生育を阻害する。光合成生物は、光を利用して、損傷した光合成装置を迅速に分解・修復する機構を持っており、それによって光合成活性を維持している。↑
注6 特別天然記念物「阿寒湖のマリモ」
阿寒湖は、阿寒カルデラおよび古阿寒湖の成立後に雄阿寒岳の噴火に伴って分断・形成された堰止湖である。阿寒湖の北に位置するチュウルイ・キネンタンペ両湾には、世界でも珍しい球状集合体のマリモが群生する。直径30 cm以上の巨大なマリモが確認された湖は世界中で阿寒湖だけである。その希少性と学術的な価値の高さから1921年に国の天然記念物、1952年に特別天然記念物に指定された。現在、国の天然記念物に指定されている藻類は8種、特別天然記念物に指定されている藻類は阿寒湖のマリモ1種である。
")
