ゲノムDNAが倍加していく様子を1細胞レベルで明らかに
2019-02-26 理化学研究所,三重大学
理化学研究所(理研)生命機能科学研究センター発生エピジェネティクス研究チームの平谷伊智朗チームリーダー、高橋沙央里基礎科学特別研究員、三浦尚研究員、三重大学大学院医学系研究科の竹林慎一郎講師、柴田隆豊大学院生(研究当時)らの共同研究グループ※は、増殖中の細胞でゲノムDNAが倍加していく過程を網羅的に解析する「scRepli-seq法」を開発し、その様子を1細胞レベルで詳しく観察することに成功しました。
真核生物の「ゲノムDNA複製[1]」の真の姿を捉えた本成果は、細胞核内でゲノムDNA複製が制御される仕組みや、クロマチン[2]高次構造とゲノム機能の関係性の理解に大きく貢献すると期待できます。
増殖中の細胞におけるゲノムDNA複製のプロセスは、これまでは技術的な限界から、数万個に及ぶ細胞の平均像としてしか捉えられていませんでした。
今回、共同研究グループは、1細胞での全ゲノム解析[3]を実現するscRepli-seq法を開発し、さらに「一塩基多型[4]」の情報を用いることで、一つ一つの細胞で父方と母方由来の染色体1本1本が複製されていく様子を捉えることに成功しました。染色体のどの領域がいつ複製されたかを集団中の個々の細胞間で比較すると、そのゆらぎ(ばらつき)は予想以上に小さく、ゲノムDNA複製の時間的制御は、細胞間で高度に保存されていました。つまり、これまで見えていた数万個の細胞の平均像は、個々の細胞におけるゲノムDNA複製の姿に驚くほど近かったことが明らかになりました。
本研究成果は、国際科学雑誌『Nature Genetics』オンライン版(2月25日付け:日本時間2月26日)に掲載されます。

図 マウスES細胞のゲノムDNAが複製されていく様子と細胞間の複製のゆらぎ
※共同研究グループ
理化学研究所生命機能科学研究センター 発生エピジェネティクス研究チーム
チームリーダー 平谷 伊智朗(ひらたに いちろう)
基礎科学特別研究員 高橋 沙央里(たかはし さおり)
研究員 三浦 尚(みうら ひさし)
(以上、旧所属は多細胞システム形成研究センター 発生エピジェネティクス研究チーム)
三重大学大学院 医学系研究科 基礎医学系講座 機能プロテオミクス分野
教授 緒方 正人(おがた まさと)
講師 竹林 慎一郎(たけばやし しんいちろう)
三重大学大学院 生物資源学研究科 生命機能化学講座 分子細胞生物学分野
教授 奥村 克純(おくむら かつずみ)
大学院生(研究当時)柴田 隆豊(しばた たかひろ)
(現 滋賀県農業技術振興センター)
大阪大学大学院 理学研究科 生物科学専攻 染色体構造機能学研究室
教授 小布施 力史(おぶせ ちかし)
准教授 長尾 恒治(ながお こうじ)
※研究支援
本研究は、日本学術振興会(JSPS)科学研究費補助金若手研究「1細胞DNA複製タイミング解析による発生分化過程の核内コンパートメント動態予測(研究代表者:高橋沙央里)」、同新学術領域研究(研究領域提案型)「染色体機能ドメインの可塑性とその意義(研究代表者:竹林慎一郎)」、同新学術領域研究(研究領域提案型)「細胞分化にともなうクロマチンポテンシャルの変化とその分子基盤(研究分担者:平谷伊智朗)」、同新学術領域研究(研究領域提案型)「ヘテロクロマチンの凝縮構造を作り出すノンコーディングRNA群の解明(研究代表者:長尾恒治)」、同新学術領域研究(研究領域提案型)「性染色体ヘテロクロマチンが規定する性スペクトラム(研究代表者:長尾恒治)」による支援を受けて行われました。
背景
生物が持つ基本的な性質の一つは、「増える」ことです。あらゆる生物の細胞は、生命の設計図であるゲノムを正確に倍加させる「ゲノムDNA複製」というプロセスを経て初めて、二つの娘細胞に分かれ、増殖することができます。
真核生物では、細胞周期[5]のS期[5]にゲノムDNAが複製される際に、どの領域から順に複製が進行していくかという順序(複製タイミング[6])と、それらの領域におけるクロマチン高次構造との間に強い相関が見られることが報告されています注1)。すなわち、DNAが緩く折り畳まれて遺伝子がよく転写[7]されている領域ほど複製タイミングが早く、密に折り畳まれて遺伝子の転写が抑えられている領域ほど複製タイミングが遅いことが知られています。
複製の過程を正確に理解することは、細胞分裂を通してゲノムDNAが安定に維持され、クロマチン高次構造が適切に制御される仕組みの理解という観点から非常に重要です。しかし、多細胞生物を構成する一つ一つの細胞について、複製タイミングとクロマチン高次構造がどの程度関連しているかは分かっていませんでした。複製の過程を網羅的に調べるには、少なくとも数万個の細胞が必要であり、そのような解析から得られた複製に関する記述は、実は数万個の細胞から得られた平均像にすぎなかったからです。個々の細胞で実際にどのように複製が進行するのかは、分子生物学における未解決の問題でした。
注1)Rhind, N.and Gilbert, D.M. DNA replication timing. Cold Spring Harb Perspect Biol. 5, 1-26 (2013).
研究手法と成果
1細胞レベルで解析を行うためには、従来法とは根本的に違う方法論が必要です。共同研究グループは、S期の細胞1個からゲノムDNAを取り出し、各領域間のコピー数の差を検出できれば、複製前(1コピー)と複製後(2コピー)の領域を識別できる可能性があると考えました。この考えをもとに新しい方法論の開発に取り組み、ゲノムDNAが複製される様子を1細胞レベルの全ゲノム解析によって捉える「scRepli-seq(single-cell DNA Replication sequencing)法」の開発に成功しました。この手法でS期中期のヒト細胞を解析した結果、全ての染色体について、どの領域がどこまで複製されているかを詳細に表す「DNA複製プロファイル」が得られました(図1)。
さらに共同研究グループは、scRepli-seq法を用いた全ゲノム解析に、もう一つ工夫を加えました。父方由来と母方由来の相同染色体[8]はほぼ同じDNA配列を持ちますが、父母間で異なる「一塩基多型」の情報を利用することで、1細胞中の各染色体について父方由来と母方由来の染色体を識別することができます。scRepli-seq法にこの識別を組み合わせることで、細胞中の染色体1本1本がまさに複製されていく様子を捉えることにも成功しました(図2)。
さらに、マウスES細胞[9]を解析した結果、1細胞ゲノムDNA複製プロファイル間のゆらぎ(ばらつき)は、予想よりもはるかに小さく、複製のプロセスは細胞間で高度に保存されていました(図3)。また、マウスES細胞を分化[10]させたところ、分化後にDNA複製プロファイルは大きく変わるものの、分化後も細胞集団中のゆらぎは依然小さいことも分かりました(図3)。これらの細胞において、相同染色体同士はよく似たプロファイルを示したことから、特定の分化状態の均一な細胞集団において、複製プロセスの時間的制御は、細胞間のみならず、相同染色体間でもかなり保存されていることが明らかになりました(図2)。
以上の結果から、これまで数万個の細胞の平均像として見えていたゲノムDNA複製のプロセスは、個々の細胞のそれにかなり近いことが明らかになりました。
次に、複製タイミングとクロマチン高次構造の相関について、1細胞レベルで検証することを試みました。近年のHi-C法[11]と呼ばれる全ゲノム解析を用いた染色体の三次元構造の研究から、間期[5]の細胞核の中で各々の染色体は100万塩基対[12]ほどの球状に折り畳まれたトポロジカルドメイン(TAD:Topologically Associating Domain)が数珠つながりになった構造をとると考えられています(図4上)。TADはさらに折り畳まれ、遺伝子がよく転写されているTADが集まってAコンパートメントと呼ばれる領域を、転写されていないTADが集まってBコンパートメントと呼ばれる領域を核内空間に形成するとされています注2)。
scRepli-seq法によって得られた1細胞DNA複製プロファイルは、このA/Bコンパートメントの分布を非常によく反映していました(図4下)。このことは、A/Bコンパートメントという構造の分布も、細胞間で高度に保存されている可能性を示唆しています。
注2)Lieberman-Aiden, E. et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science (New York, N.Y.) 326, 289–93 (2009).
以上の結果から、均一な細胞集団に属する個々の細胞は、よく似たDNA複製プロファイルを持つという傾向が明らかとなりました。一方で、DNA複製プロファイルの中で差異の大きい箇所、すなわち細胞間・相同染色体間で比較的ゆらぎの大きい領域を詳しく見ていくと、以下の特徴があることも分かりました。
- 細胞間の複製時期のゆらぎは、S期の初期と後期でかなり小さく、中期に複製される領域についてはゆらぎが比較的大きい(図5)。
- 細胞分化[10]に伴い複製タイミングが変化する領域に注目すると、未分化ES細胞の段階で既に細胞間の複製時期のゆらぎが比較的大きい。
これらの観察結果は、①ゲノムDNAの複製がどの領域から開始してどの領域で完了するかは、細胞の種類ごとにあらかじめほぼ決定されているが、それ以外の領域がいつ複製されるかは細胞間で多少ゆらぐ、②細胞分化に伴う複製タイミングの変化は、その変化に先立つ複製時期のゆらぎ、すなわちコンパートメント構造の不安定性に起因している可能性がある、と解釈できます。これらの結果は、1細胞ゲノムDNA複製プロファイルを解析して初めて明らかになった現象であり、いまだにほぼ手付かずの状態にある複製タイミングおよびA/Bコンパートメント構造の制御メカニズムを考える上で重要な発見です。
今後の期待
本研究で、共同研究グループは、ゲノムDNA複製の時間的制御が個々の細胞間でほぼ共通であることを明らかにしました。scRepli-seq法は非常にシンプルで汎用性が高く、あらゆる生物種に適用できるため、今後、DNA複製研究に大きく貢献することは確実です。一方、複製時期が細胞間で比較的大きくゆらぐ領域の存在も明らかになりました。このゆらぎは、ゲノムDNAの局所的なクロマチン構造のゆらぎと考えられ、クロマチン構造制御に関する新たな問いを生み出すものでもあります。
scRepli-seq法の確立により、ゲノムレベルのDNA複製研究、そしてクロマチン構造研究の一部については、細胞数のハードルはもはや完全に取り払われたことになります。今後は本手法を活用して、個体や臓器中の個々の細胞におけるゲノムDNAの振る舞いや状態を理解する研究が進み、複製異常や染色体構造異常を伴うさまざまな疾患の理解にもつながることが期待できます。
原論文情報
Saori Takahashi, Hisashi Miura, Takahiro Shibata, Koji Nagao, Katsuzumi Okumura, Masato Ogata, Chikashi Obuse, Shin-ichiro Takebayashi, Ichiro Hiratani, “Genome-wide stability of the DNA replication program in single mammalian cells”, Nature Genetics, 10.1038/s41588-019-0347-5
発表者
理化学研究所
生命機能科学研究センター 発生エピジェネティクス研究チーム
チームリーダー 平谷 伊智朗(ひらたに いちろう)
基礎科学特別研究員 高橋 沙央里(たかはし さおり)
研究員 三浦 尚(みうら ひさし)
三重大学大学院 医学系研究科 基礎医学系講座 機能プロテオミクス分野
講師 竹林 慎一郎(たけばやし しんいちろう)
大学院生(研究当時) 柴田 隆豊(しばた たかひろ)
お問い合わせ先
理化学研究所 生命機能科学研究センター センター長室 報道担当
山岸 敦(やまぎし あつし)
報道担当
理化学研究所 広報室 報道担当
補足説明
-
- ゲノムDNA複製
- 細胞分裂における核分裂の前に、DNAポリメラーゼという酵素によって、ゲノムDNAが過不足なく倍加される過程。ゲノム複製、DNA複製と同義。ゲノムDNAとは、生物の細胞が持っている全てのDNA配列情報のことを指し、真核生物の場合は核内にある染色体DNAを指すことが多い。
-
- クロマチン
- 真核生物の核内で、ゲノムDNAが形成する高次構造体のこと。その主成分は、DNAとヒストンと呼ばれるタンパク質。
-
- 全ゲノム解析
- ヒトやマウスなど、単一の生物種のゲノムを構成する全てのDNA配列にわたって、特定の性質(RNA転写量、特定のDNA結合タンパク質の結合性など)を、次世代シーケンサーを用いて網羅的に調べる解析のこと。
-
- 一塩基多型
- ある生物種集団のゲノムDNA配列中に1塩基が変異した多様性が見られ、その変異が集団内で1%以上の頻度で見られるとき、これを一塩基多型(SNP; Single Nucleotide Polymorphism)という。塩基とは核酸の構成要素のことで、DNAの場合はアデニン(A)、チミン(T)、グアニン(G)、シトシン(C)の4種類を指す。実際には、1塩基に限らず、2塩基から十数塩基程度の短い置換、挿入、欠失などの変異も含めて一塩基多型と呼ぶことも多く、本研究でもこの立場をとっている。
-
- 細胞周期、S期、間期
- 増殖中の真核細胞が繰り返す時間的なサイクル(期間)のことを細胞周期と呼ぶ。間期とは、細胞周期の中で細胞分裂を行う分裂期(M期)以外の期間のこと。間期は、さらにG1期、S期、G2期に分けられる。S期は、DNA複製が行われる期間。
-
- 複製タイミング
- ゲノムDNA複製の時間的制御のこと。S期において、各ゲノムDNA領域は、それぞれ特徴的な時期に複製されると考えられている。本研究以前は、その時間的制御が1細胞レベルで、どの程度ゆらぐのかは未解決の問題であった。
-
- 転写
- 染色体DNAの遺伝子配列をもとにRNAが合成されること。その担い手はRNAポリメラーゼと呼ばれる酵素。
-
- 相同染色体
- 二倍体の生物の細胞核内に存在する、父方および母方から由来した形態の等しい一対の染色体のことを指す。ヒトの細胞は、1番から22番染色体まで22対の相同染色体を持ち、これに加えて2本の性染色体を持つ(オスはXY、メスはXX)。
-
- ES細胞
- 胚性幹細胞(Embryonic Stem Cell; ES cell)とも呼ばれる。哺乳類生物の発生初期の胚盤胞期と呼ばれる時期の胚の一部、内部細胞塊と呼ばれる細胞群から作られる幹細胞細胞株のこと。性質はiPS細胞(人工多能性幹細胞; induced Pluripotent Stem Cell)とよく似ているが、iPS細胞は体細胞から作られる細胞株である。
-
- 分化、細胞分化
- 分化ともいう。多細胞生物の発生過程において、特殊化していない未分化の細胞が、より特殊化したタイプの細胞に変化するプロセスのことを指す。
-
- Hi-C法
- 3C(Chromosome Conformation Capture)法を発展させた全ゲノム解析手法のことで、細胞核内三次元空間におけるあらゆるゲノムDNA配列どうしの相対距離を測定して、染色体の三次元的な構造を推定できる画期的な手法。
-
- 塩基対
- ゲノム中で2本のデオキシリボ核酸(DNA)分子が2重らせん構造をとる際に、アデニン(A)とチミン(T)、グアニン(G)とシトシン(C)、という決まった組を作り、水素結合でつながった状態をとるが、その決まった各々の組のことを塩基対と呼ぶ。DNAの長さを表現するときに「~塩基対(base pairs: bp)のDNA」、あるいは単に「~塩基(bases : b)」という表現が用いられることが多い。Mbはメガベース、100万塩基(対)のこと。Kbはキロベース、1000塩基(対)のこと。
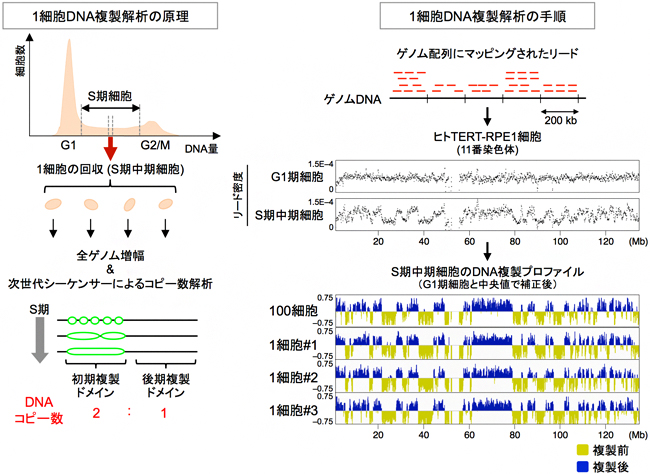
図1 本研究で開発した1細胞ゲノムDNA複製解析法「scRepli-seq法」の原理と解析の手順
左: scRepli-seq法の原理(図はS期中期細胞の場合)。フローサイトメーターという細胞を分ける機器を用いて、S期の細胞を一つ一つ個別に回収する。各々の細胞から抽出したゲノムDNAの全ゲノム増幅を行い、次世代シーケンサーを用いてこのゲノムDNAを解読し、ゲノムDNA上の初期複製ドメイン(複製後)と後期複製ドメイン(複製前)をDNAコピー数によって識別する。
右: 解析の手順。(上段)次世代シーケンサーで得られた配列(リード)をマッピング(配列が一致するゲノム領域にリードを対応させる作業)し、解析を行った。(中下段)ヒトTERT-RPE1細胞(網膜色素上皮細胞)の11番染色体のデータで、横軸に染色体上の位置と距離を示している(Mb:メガベース、100万塩基対)。染色体ごとに見ると、複製前のG1期の細胞では、染色体全域でリード密度がほぼ一定なのに対し、S期中期の細胞ではリード密度が領域によって異なることが分かる(中段)。S期中期細胞のリード密度を、G1期細胞データ及びS期中期細胞の中央値で補正すると、複製前(黄)と複製後(青)の領域が存在すること、また、1細胞での結果が100細胞からの結果がほぼ同じであることが分かる(下段)。

図2 父方由来と母方由来の染色体を識別したscRepli-seqデータ
左:解析の流れ。母=CBA系統(西ヨーロッパ産マウス亜種由来)、父=MsM系統(日本産マウス亜種由来)の掛け合わせで生まれた雑種第一代(CBMS1)雌マウスに由来するES細胞を用い、通常のscRepli-seq実験を行った後、一塩基多型の解析を行って、母方(CBA)由来と父方(MsM)由来の染色体を識別し、最終的にそれぞれの染色体のDNA複製プロファイルを完成させる。
右:解析結果。CBMS1マウスES細胞について、合計125個の1細胞データからの母方(CBA)由来と父方(MsM)由来の染色体ごとの1細胞DNA複製プロファイルを示す。細胞は、S期全域から回収し、ゲノムDNA複製率(ほとんど未複製なものから、複製完了に近いものまで)に従って並べた。1万細胞の平均像として見えていたゲノムDNA複製のプロセスが、個々の細胞のそれにかなり近いことが分かる。

図3 分化前のES細胞と分化後の細胞のscRepli-seqプロファイルのゆらぎ
tSNE法というデータ解析手法を用いて、105個の細胞のDNA複製プロファイルのゆらぎ(ばらつき)を2次元のグラフで表現した。解析に用いたのはS期中期にあるES細胞(48個)と、ES細胞から分化させた細胞(42個)で、G1期細胞は対照群(ばらつきが少なくグラフ上でまとまった集団)。個々の細胞間に多少のばらつきはあるものの、ES細胞、分化細胞、G1期細胞はそれぞれ違う個性を持ち、同一細胞種内での1細胞DNA複製プロファイルのゆらぎは比較的小さいことが分かった。

図4 A/Bコンパートメント分布と1細胞レベルの複製ドメイン分布の相関
上段は、細胞核内の染色体三次元構造の模式図と、マウスES細胞のHi-C解析によって得られたA/Bコンパートメント分布を示す。下段にはS期中期細胞から得られたscRepli-seqデータを示す。「由来の区別なし」は、父方母方由来染色体識別前のデータ。「母方由来」と「父方由来」は識別後のデータ。1細胞ごとのゲノムDNA複製プロファイルは、細胞集団の解析から得られたA/Bコンパートメント分布に非常によく似ており、上段のAコンパートメント(緑)とBコンパートメント(赤)は、それぞれ青色の複製後(S期前半に複製される領域)と黄色の複製前(=S期後半に複製される領域:黄)のゲノムDNA領域に対応していた。

図5 S期初期・後期および中期におけるゲノムDNA複製時期のゆらぎの大きさ
S期全域から得られたマウスES細胞の1細胞DNA複製解析データのうち、14番染色体の一部を示す。下の緑の棒グラフは、ゲノムDNAのある領域を複製し終えた細胞の割合が25%から75%になるのに要する時間をT25->75と定義し、各領域ごとの時間を示したもの。T25->75が大きいほど細胞間の複製時期のゆらぎは大きい。S期初期と後期に複製される領域はT25->75が非常に短く、対照的に、S期中期に複製される領域はT25->75が長い。

