2022-12-02 東京大学

発表者
上浦 六十(東京大学 大学院総合文化研究科 広域科学専攻 博士課程)
水内 良(東京大学 大学院総合文化研究科 附属先進科学研究機構 特任助教/科学技術振興機構 さきがけ研究者)
市橋 伯一(東京大学 大学院総合文化研究科 広域科学専攻・附属先進科学研究機構 教授/東京大学 生物普遍性連携研究機構 教授)
発表のポイント
- 生命の起源において、自己複製する単純な分子から最初の生命が生まれたと想像されているが、単純な分子がどのようにして複雑な生命体へと進化したのかはわかっていなかった。
- 自己複製RNAを用いた実験と計算機シミュレーションを用いた理論解析の両面から、寄生型の自己複製体の出現が複製ネットワークの複雑化を可能にしたことを示す証拠を得た。
- 寄生体を介した複製ネットワークの複雑化が、多機能で複雑な生命システムへ至る進化の第一歩となった可能性を見出した。
発表概要
東京大学大学院総合文化研究科広域科学専攻の上浦六十大学院生、同専攻・附属先進科学研究機構/東京大学生物普遍性機構の市橋伯一教授らの研究グループは、単純な自己複製分子が複雑性を進化させていく過程に寄生型分子との相互作用が重要だった可能性を、実験と計算機シミュレーションの両面により明らかにしました。
原始地球において、単純な自己複製能力を持つ分子(RNAワールドにおける自己複製RNAなど)がダーウィン進化(注1)する能力を獲得し、この能力により次第に複雑性を増していくことで最初の生命体が生まれたと想像されています。しかし、ダーウィン進化の仕組みで複雑化していくことは容易なことではありません。これまでに行われた自己複製分子を使った進化実験では、多くの場合、増える以外の機能をそぎ落とした最も単純な分子が進化してしまい、複雑化した分子が進化したことはありませんでした。それではどのようにして、単純な自己複製分子が複雑性を進化させることができるのでしょうか?これは生命の初期進化過程における大きな謎として残っています。
これまで本研究グループは、RNAとタンパク質からなるRNAの自己複製システムを用いて長期の実験室内進化実験を行い、単純な自己複製RNAの進化過程を研究してきました。そのなかで自己複製RNAから寄生型のRNAが出現すること、そして、この寄生型RNAと元の宿主RNAが共進化し最大5種類のRNAが相互作用しながら複製する複雑な複製ネットワークが出現することを見出しました。本研究では、なぜこのような複雑なネットワークが進化することができたのかを明らかにするために、数理モデルと計算機シミュレーションを用いたネットワークの安定性解析と検証実験を行いました。その結果、先行する理論研究で提唱されているように、寄生型のRNAがいわゆるニッチ (注2)として働くことで複数の宿主RNAが共存を可能にしていることを明らかにしました。この結果から、寄生体を介した複雑化進化が単純な分子から複雑な生命システムへといたる進化過程の第一歩になったのではないかと考えています。
本研究成果は、2022年12月1日(米国東部時間)に米国科学誌「Plos Computational Biology」のオンライン版に掲載されました。
発表内容
生命が生まれる前の時代には、RNAや短いタンパク質などの分子が集まってできた単純な自己複製分子が存在しており、それらが少しずつ進化して複雑化していった結果、私たちすべての生物の祖先(細菌のような単細胞だと考えられています)が誕生したと想像されています。しかし、単純な自己複製分子が進化して複雑化していく様子を観察することは、いままで誰もできていません。むしろ実験室で自己複製分子を進化させた場合、増えることに必要な機能以外を失って単純化することが知られています。例えば、アメリカのシュピーゲルマンらのグループは1967年に初めて試験管内で進化する自己複製RNAを報告しましたが、そのRNAはどんどん短くなり単純化していきました。これは単純なRNAの方が少ない材料で速く複製することができるためです。いったいどのようにして単純な自己複製分子が複雑性を進化させることができるのでしょうか?これは生命の起源に関わる大きな謎として残されていました。
この謎を解明するために、本研究グループはこれまでに独自に開発したRNAの自己複製システム(図1)を用いて長期の実験室内進化実験を行ってきました。そのなかで自己複製RNAから、複製酵素遺伝子を持たず宿主が作った複製酵素に依存して増える寄生型のRNA(寄生体)が出現すること(図2)、そして、この寄生型RNAと元の宿主RNAが共進化し、最大5種類のRNAが相互作用しながら複製する複雑な複製ネットワークが進化することを見出しました(図3)。しかし、なぜ本研究グループの実験ではシュピーゲルマンの実験とは違い自己複製RNAが複雑性を進化させることができたのか、その理由はあきらかではありませんでした。過去の理論研究から、自己複製RNAに寄生型のRNAが生まれると、寄生型がニッチとして機能することで複数の自己複製RNAが共存できることが提唱されていました。そこで本研究では、この仮説が本研究グループの実験にも成り立っているかどうかを数理モデルと計算機シミュレーションによるネットワークの安定性解析と検証実験により明らかにすることを試みました。
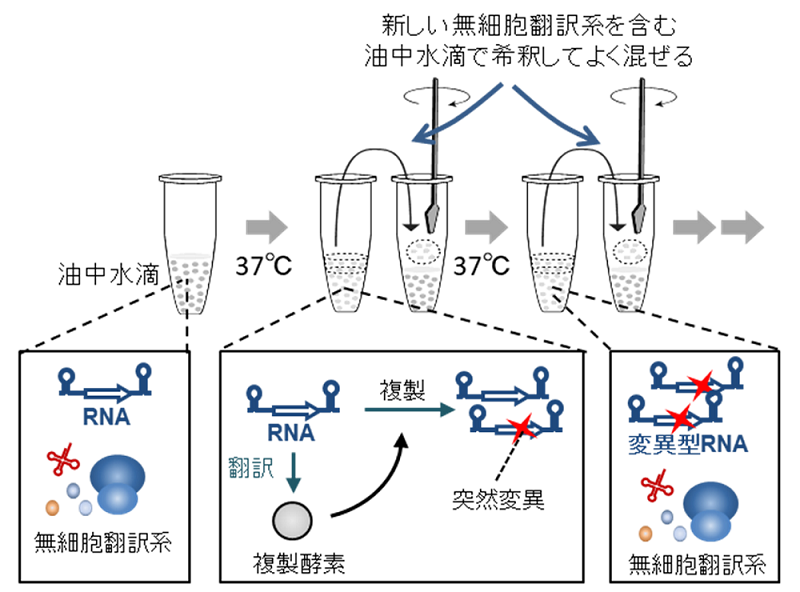
図1.RNAの自己複製システムを用いたRNAの進化実験の模式図
本システムを微小油中水滴に封入し、新しい翻訳系を含む油中水滴で一定希釈しながら継代するとRNA複製を続けることができる。複製中には複製ミスにより変異が入る。もし変異の入った変異型RNAが元のRNAよりも複製能力が高ければ、より多くの子孫を残すため集団を占めることになる。これは生物が適応進化するメカニズムと同じ現象である。
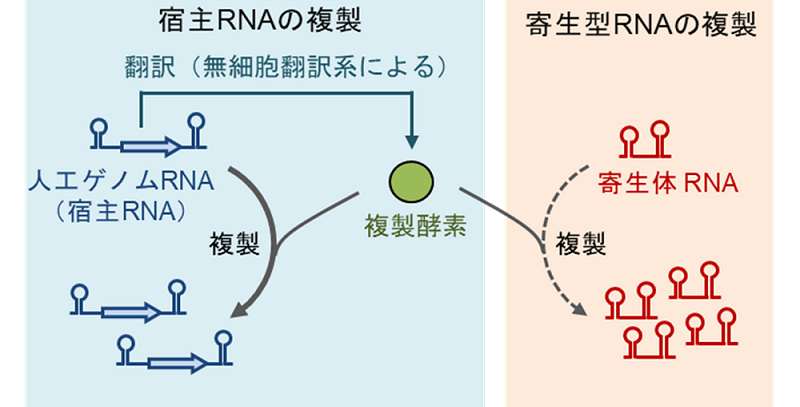
図2.宿主RNAと寄生体RNAの模式図
宿主RNAは複製酵素を作って単独で複製できるが、寄生体RNAは複製酵素遺伝子を持たないため、宿主が作った複製酵素に依存してのみ増えることができる。
(Furubayashi, T., Ueda, K., Bansho, Y., Motooka, D., Nakamura, S., Mizuuchi, R., & Ichihashi, N. (2020). Emergence and diversification of a host-parasite RNA ecosystem through Darwinian evolution. ELife, 9. https://doi.org/10.7554/eLife.56038)
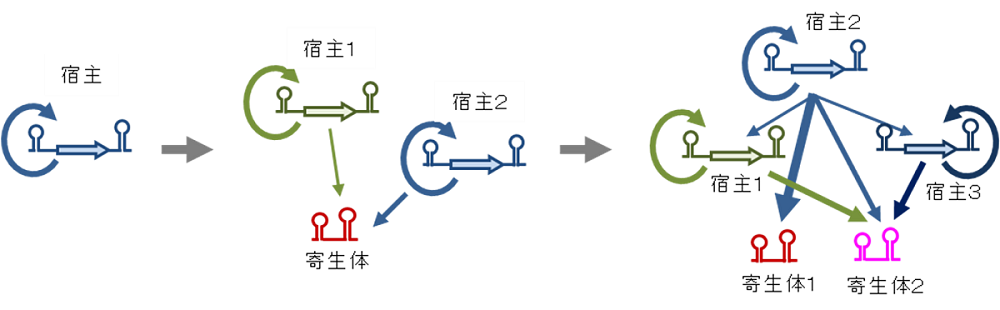
図3.複数の宿主、寄生型RNA が相互作用した複雑な複製ネットワークの出現
これまでに行った長期進化実験により、もともと単一だった宿主RNAから寄生型RNAが出現し、さらに宿主、寄生型RNAの両方が分化し、お互いに相互作用しながら複製するネットワークを形成した。
(Mizuuchi, Ryo., Furubayashi, Taro., & Ichihashi, Norikazu. (2022). Evolutionary transition from a single RNA replicator to a multiple replicator network. Nature Communications, 13, 1460. https://doi.org/10.1101/2021.09.09.459571)
まず本研究グループが用いている自己複製RNA複製システムの数理モデル化を行いました(図4)。この数理モデルを使った計算機シミュレーションにより、実験では確かめられないような膨大なパラメータの組み合わせで複製ネットワークの安定性を調べることができます。本研究では最大3種類の宿主RNA(H)と寄生型RNA(P)が構成するネットワークについて、現実的なパラメータの組み合わせを網羅的に解析し、複数種類のRNAが共存しながら複製可能なネットワークを調べ上げました。その結果、複数種類の宿主のみから形成される複製ネットワーク(HHやHHHネットワーク)では、ほとんどすべての条件で、すぐに単一種類の宿主のみになってしまうことを見出しました(図5A)。つまり、たとえ複数の異なる宿主RNAが出現したとしても、存続できる宿主RNAはそのうち最も速く増える1種類だけだということです。一方で宿主に加えて寄生体が存在すると、多くの条件で2種類の宿主RNAが共存できるようになることを見出しました(図5B)。このとき共存できた2種類の宿主RNAは、寄生型RNAに対して異なる耐性を持ち、かつ耐性度の高い宿主RNAが低い宿主RNAをよく増やすような性質を持っている傾向がありました。この結果は、宿主RNAと寄生型RNAが相互作用する複製ネットワークにおいて、寄生体の存在によって複数の異なる性質を持つ宿主が共存できるようになることを示しています。
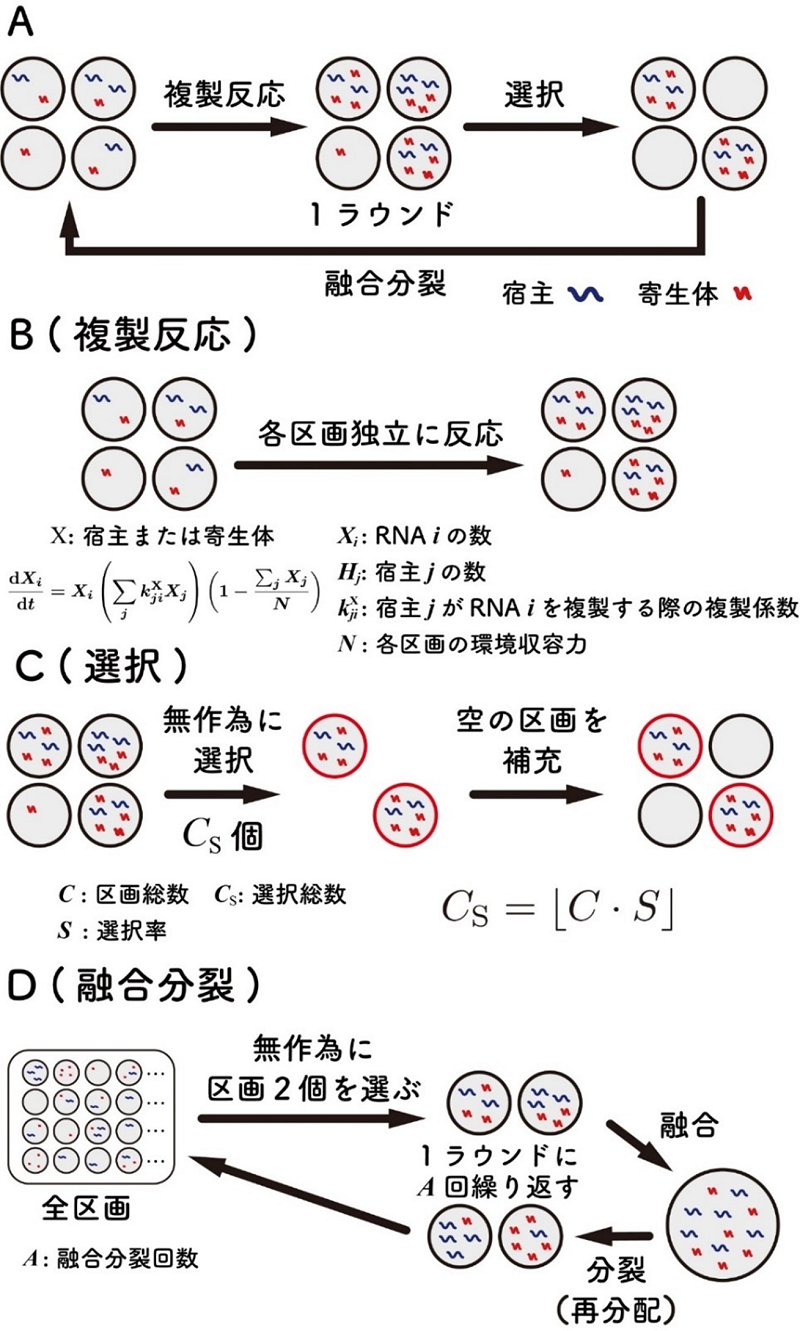
図4.自己複製RNA複製システムの数理モデル化
図1に示す進化実験を図中AからDのステップによりモデル化し計算機シミュレーションを行った。
(A)計算機シミュレーションの全体像。このシミュレーションは微小区画構造中の複製ステップ、区画の選択ステップ、区画の融合と分裂ステップの3ステップからなる。
(B)複製ステップでは宿主と寄生型RNAが各区画内でBに示す微分方程式に従って複製する。
(C)選択ステップでは、ある数(Cs)の区画がランダムに選択され、それ以外の区画はRNAの入っていない空の区画で置き換えられる。
(D)融合分裂ステップでは2つの区画をランダムに選び、その中身の宿主と寄生型RNAを混ぜ、その後、2つの区画にランダムに分配した。この融合分裂ステップを5,000回繰り返した。区画の数は3,000とした。
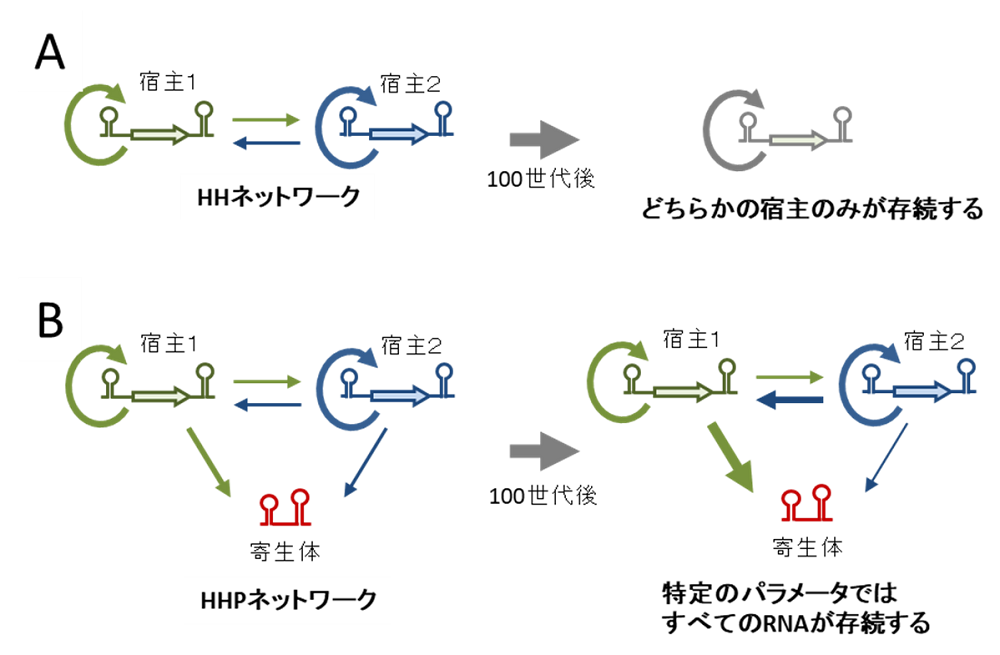
図5.寄生体の存在により2種類の宿主RNAの共存が可能になる
複製ネットワークの名称は宿主RNA(Host, H)と寄生型RNA(Parasite, P)の組み合わせで表現されている。例えばHHネットワークは2種類の宿主RNA(H)からなり、HHPネットワークは2種類の宿主(H)と1種類の寄生型RNA(P)からなる。
(A)2種類の宿主からなる複製ネットワーク(HHネットワーク)では、ほとんどのパラメータ(複製係数)で100世代後にはどちらか単一の宿主のみになってしまった。
(B)加えて1種類の寄生型RNAがいる場合(HHPネットワーク)では多くのパラメータで2種類の宿主RNAと1種類の寄生型RNAが共存できた。この時、宿主1と2には寄生型RNAに対して異なる耐性度を持っており、耐性度の高い宿主がそうでない宿主をよく複製する傾向があった。
以上の結果から、自己複製分子の複製ネットワークが複雑性を進化させる経路には一定のルールがあることが予想されます。つまり、最初の宿主自己複製分子が出現すると、次に共存できるのは別の宿主分子ではなく、寄生型の自己複製分子 となります。こうしてHPネットワークが形成されます。次に共存できるのは、その寄生体に対し耐性を持つ別の宿主分子です。こうしてHHPネットワークが形成されます。このようにして、さらに次は別の寄生体、次にそれに耐性を持つ 別の宿主といったように、寄生体とそれに耐性を持つ宿主が順番に追加されていくことで、共存状態を維持したまま複製ネットワークが複雑化することが可能となると予想されます(図6)。また本研究では、パラメータが次々に変化する進化シミュレーションによっても確かにHPネットワークからHHPネットワークへの変遷が起こることを確かめています(図7)。
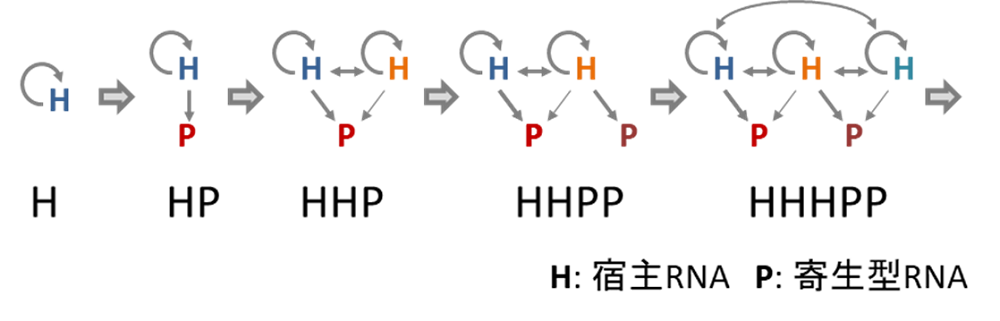
図6.自己複製ネットワークが複雑化する際のルール
網羅的なシミュレーションの結果から、宿主RNAと寄生型RNAが順番にネットワークに追加されていくと複数のRNAが共存したままネットワークが複雑化できる。
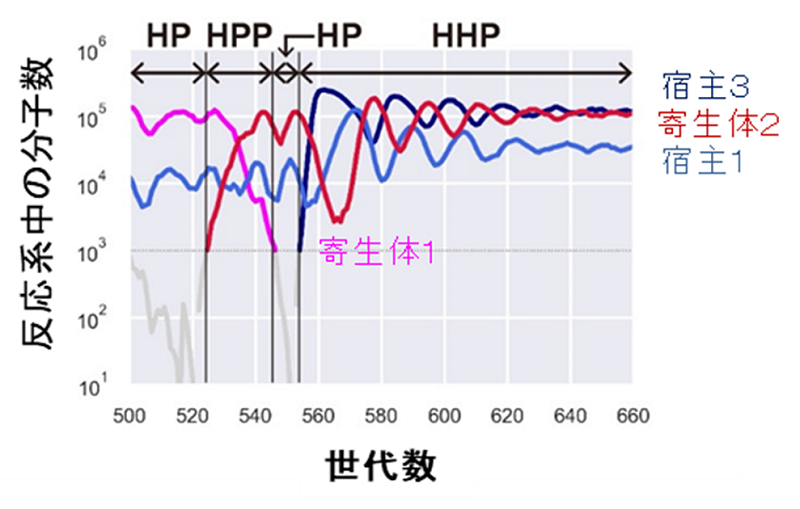
図7.変異によりパラメータが変化する場合の進化シミュレーション
一定頻度でランダムなパラメータを持つ宿主あるいは寄生型RNAが出現する条件で進化シミュレーションを行った。まず宿主1種類(宿主1)と寄生体1種類(寄生体1)のHPネットワークから寄生体が2種類に増えたHPPネットワークが生まれたが、片方の寄生体(寄生体1)がすぐに共存できなくなり、HPネットワークに戻った。その後、別種の宿主(宿主3)が生まれ、安定したHHPネットワークを形成した。このシミュレーションでは、計算量を抑えるためにある時間に出現可能な最大のRNAの種類を3以下に限定している。
さらに、これまで本研究グループが行ってきた自己複製RNAの進化実験中に、このルールに従って進化が起きているかを確かめました。進化実験は単一種類の宿主RNAから開始しましたが、すぐに寄生型RNAが出現し、その後、宿主RNAは2系統へと分岐していきました。各系統の代表的なRNAを単離しその複製パラメータ (どのRNAがどのRNAをどのくらい増やすかを示す値)を測定したところ、確かに安定した HPおよびHHPネットワークを形成するパラメータとなっており、進化実験中にもHPからHHPネットワークの変遷が起きていたことが支持されました。
本研究の結果から、原始の自己複製分子は寄生型分子の出現を介して複雑性を進化させていったことが考えられます。つまり、寄生体がいない状態での宿主どうしの競争ではどちらか少しでも複製が速い方しか存続することができませんが、寄生体の出現により、寄生体に対する抵抗性の違いにより複数の宿主が共存できるようになり、それらが相互作用することで複雑な複製システムが生まれたということです。これまでの進化実験ではHHHPPの5者ネットワークまで出現しています(図3)。これら5種類のRNAはもともとは単一のRNAでしたが、継代反応を繰り返すうちに少しずつ異なる機能(特定のRNAの複製しやすさとされやすさ)を持ち、相互作用しながら複製するように進化しました(図1)。この相互作用しながら複製する分子ネットワークは、過去にマンフレッド・アイゲンが提唱したハイパーサイクル(注3)によく似ています。本研究グループは、この相互依存する5種類のRNAが今後、1種類の長いRNAへと融合していくのではないかと予想しています。このようにして分化と融合を繰り返すことで原始自己複製体は次第に多くの機能を獲得し、複雑な生命体へと近づいて行ったのではないかと想像しています。このような寄生体を介した複雑化進化が、単純な分子から複雑な生命システムへといたる進化過程の第一歩になったのではないかと考えています。
論文情報
雑誌:Plos Computational Biology(オンライン版:2022年12月1日掲載)
論文タイトル:Plausible pathway for a host-parasite molecular replication network to increase its complexity through Darwinian evolution
著者:Rikuto Kamiura, Ryo Mizuuchi, and Norikazu Ichihashi*
DOI番号:10.1371/journal.pcbi.1010709
用語説明
(注1)ダーウィン進化
ダーウィンが述べた多様性と自然選択により、個体、あるいは分子集団の中でより多くの子孫(分子の場合は自身のコピー)を残す個体(あるいは分子)が集団内での割合を増やしていく過程を指す。この過程を繰り返すことによって、その集団を占める個体(あるいは分子)はより多くの子孫(あるいはコピー)を残すように変化していく。
(注2)ニッチ
餌や住処など生態的な地位のことを指す。例えば、食べる餌や住む場所が異なる生物は「異なるニッチを占める」などと表現する。本研究の場合は、寄生型のRNAが存在するおかげで別のタイプの宿主RNAが共存できるようになったため、「寄生型のRNAがいわゆるニッチとして働く」と表現している。
(注3)ハイパーサイクル
最初の生命へ至る前の原始自己複製分子のモデルとして、マンフレッド・アイゲンが提唱したモデル。複数の自己複製分子が相互依存しながら全体として複製するシステム。相互依存することによって、エラーの多い原始的な複製様式でもエラーカタストロフを防いで多くの情報を持つことができる。本研究グループが見出した複製ネットワークでは、普通のハイパーサイクルのように完全な相互依存性があるわけではなく、相互依存性が直接的ではないことから、緩いハイパーサイクルとなっている。

