2023-07-20 秋田県立大学,甲南大学,基礎生物学研究所
植物は、重力や光などの環境変化に応じて自身の身体を曲げますが、同時に、曲げ過ぎないようにブレーキをかけていることは、意外と知られていません。植物が重力に逆らって茎を曲げる現象は重力屈性*1とよばれ、1880年のダーウィン親子による著書にも取り上げられているように、多くの研究者の関心を集めてきました。しかし、植物がどのように身体を曲げ、そして曲げ過ぎないように姿勢を正しているのかは、不明なままでした。
この謎に満ちた「曲げに対して姿勢を正すしくみ(すなわち,姿勢制御)」は、植物が自身の身体の状態を認識する自己受容性感覚*2の一環と捉えることができます。モータータンパク質であるミオシン*3の機能が損なわれたシロイヌナズナ変異体ではブレーキがかからず、曲がり過ぎた異常な姿勢をとることがわかっていました。
秋田県立大学、甲南大学、京都大学、奈良先端科学技術大学院大学、基礎生物学研究所、埼玉大学の共同研究グループは、変異体と比較することで、植物の姿勢を力学的な視点から調べました。茎を変形する棒とみなす弾性棒理論*4を応用し、数理力学モデルを構築しました.これによって、茎が曲がる過程で、どこにどのような力がかかるのかを定量的に示す方法論の確立に、初めて成功しました。
この力学モデルから、茎が曲がり過ぎると茎の中間領域で力学的な不均衡を起こし、不安定な姿勢になることがわかりました。すなわち、植物が姿勢正しくまっすぐに伸びることは、力学的に有利であることが示されました。本研究の成果は、ダーウィンの謎を「力学」という新たな視点から展開できる可能性を示しています。
■ 成果掲載誌
本研究成果は,国際学術誌Scientific Reportsに令和5年7月17日午前10:00(グリニッジ標準時、日本時間JST 18:00)に掲載されました。
論文タイトル:Shoot gravitropism and organ straightening cooperate to arrive at a mechanically favorable shape in Arabidopsis
(シロイヌナズナは重力屈性と器官姿勢制御の協調により力学的に有利な形状を達成する)
著者:津川暁1*,三宅唯月2,岡本圭史3,豊田正嗣4,八木宏樹2,森田(寺尾)美代5,西村いくこ2,出村拓6,上田晴子2,7*
1秋田県立大学システム科学技術学部
2甲南大学大学院自然科学研究科
3京都大学大学院理学研究科
4埼玉大学理学部
5基礎生物学研究所
6奈良先端科学技術大学院大学
7甲南大学理工学部
*責任著者
DOI: https://doi.org/10.1038/s41598-023-38069-x
■ 研究の詳細
・研究の背景
植物は重力に応答して器官内の細胞に成長速度の差を与え、偏差成長*5(へんさせいちょう)することで、根は下向きに、茎は上向きに曲がることができます。この現象は重力屈性と呼ばれており、古くは1806年にナイトが報告し、後にダーウィンが1880年に著書の中で紹介したことで、多くの関心を集めてきました。近年の植物生理学の知見から、茎では、内皮細胞でデンプンを蓄積したアミロプラスト(細胞小器官の一種)が沈降することによって、重力方向を感知します。その後、成長促進効果をもつオーキシン*6と呼ばれる植物ホルモンが偏って分布し、茎が偏差成長して曲がることがわかってきました。しかし、いったん自身を曲げる方向にアクセルを踏んだ植物は、そのままでは曲がり過ぎてしまうため、どのように姿勢を制御しているかは、長い間未解明でした。2015年に、アブラナ科のモデル植物であるシロイヌナズナを使った研究から、茎や根を含む様々な器官がぐにゃぐにゃと曲がり過ぎて、姿勢を維持できないミオシンXIやアクチンの変異体が見つかりました。これらの変異体の解析から、野生型の植物は曲り始めてから一定時間経つとブレーキをかけ、まっすぐな姿勢を取り戻すことがわかりました。本研究では、この研究をさらに発展させ、ミオシン変異体でどこにどのような力がかかって姿勢を維持できなくなるかを力学的な視点から紐解くことを目指しました。
・本研究の成果
本研究は、まっすぐな姿勢をとる野生型(図1A-C)と姿勢を維持できないミオシン変異体(図1D-F)で茎の形がどのように変化するかを、画像データを用いて分析することからスタートしました。動画では何度も観察していた変異体の曲がり過ぎる現象でしたが、この画像分析によって、どのくらいの時間で、どのくらいの角度で曲がるかを定量的に示すことが可能になりました。その結果、野生型か変異体かにかかわらず、真横に倒した植物体は、約70-80分で先端から15-20 mmの部位が曲がり始めることがわかりました。140-150分後には、野生型は姿勢を正して上に向かってまっすぐ伸びましたが,変異体は大きく曲がり過ぎた姿勢を示しました。さらに、統計検定の結果、ミオシン変異体(8個体)は野生型(9個体)と比べて曲がり始めるまでの時間が短く、曲がる位置も野生型より茎の先端に近いことが明らかになりました。
 図1: 重力刺激を与え続けたときの茎の姿勢変化を、まっすぐな姿勢の野生型(A-C)とブレーキの効かないミオシン変異体(D-F)で比較した図。
図1: 重力刺激を与え続けたときの茎の姿勢変化を、まっすぐな姿勢の野生型(A-C)とブレーキの効かないミオシン変異体(D-F)で比較した図。
(A, D)植物体を横に倒した直後 (0 min),重力刺激を与えて100分,および200分後の茎の様子。(B, E)10分ごとに4時間撮影した画像から茎の形状を抽出し,茎の角度にしたがって色分けした。(C, F)茎に沿った距離と時間による時空間角度のプロット図。
この野生型と変異体の茎の曲がり方の違いをさらに詳しく調べるために、材料力学で用いられる弾性棒モデルを構築しました(図2A,B)。このモデルでは、植物の茎を変形する棒とみなして弾性ばねによって近似し、①茎の長軸に沿った方向の引張力、②茎の長軸に対して垂直な方向の曲げ力、および③重力の三つ巴の綱引きによる力学的な安定状態を考えることにより、どこにどのような力がかかるかを解析することができます。解析の結果、野生型では曲がり始めに先端付近に生じていた大きな曲げ力を徐々に緩和してブレーキをかけているのに対し、変異体では曲がった後も曲げ力にブレーキをかけることができず曲がり過ぎてしまうことがわかりました(図2D,E)。さらに、実際の植物の観察から得られたデータに最も適合したモデルにおいて、ミオシン変異体では、曲げを生むアクセルに相当する重力感受力が弱いことがわかりました(図2C)。この結果は、ミオシン変異体でアミロプラストの運動性が低く、細胞の底面側に停留するという先行実験データとも整合していました。では、なぜ野生型ではブレーキをかけてまっすぐな姿勢をとる必要があるのでしょうか?この疑問の答えとして、何か特別な力学的な条件が必要なのではないかと考え、有限要素法*7と呼ばれる力学モデルを検討しました。
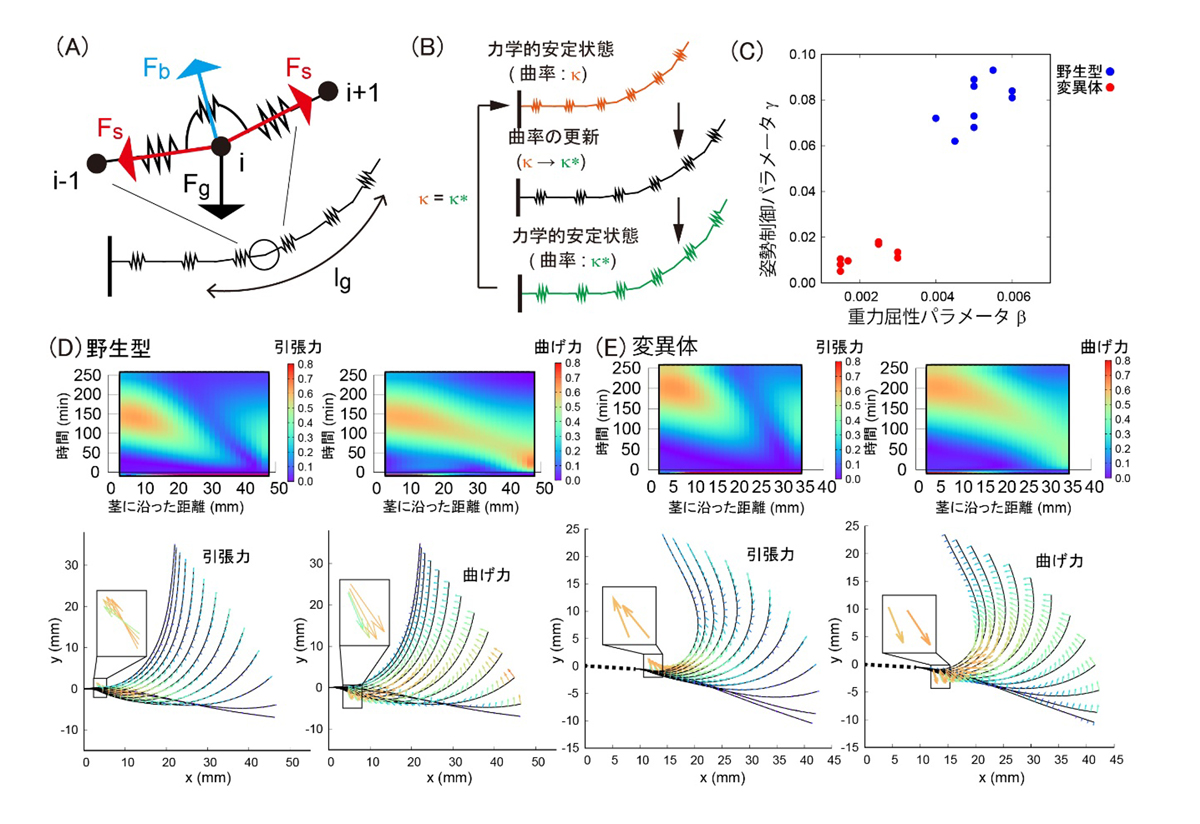 図2: 茎の重力屈性を表現する数理モデルの概要。
図2: 茎の重力屈性を表現する数理モデルの概要。
(A)茎を弾性棒とみなし、①軸方向の力、②軸外方向の力、③重力の三者で力の釣り合いを考える。(B)ある時刻の力学的安定状態を計算し、曲率を更新することで次の形を決定する。(C)実測データに最も近いモデルにおける重力屈性パラメータと姿勢制御パラメータのプロット。(D,E)野生型(D)とミオシン変異体(E)の茎の屈曲中にかかる引張力と曲げ力の解析結果。
屈曲の最中に植物体の内部でどのような力が生じるかを調べるために、有限要素法による力学モデルを再構築しました。このモデルによって、植物体の形と植物体自身の重さの関係で決まる内部応力が、どのように分布するかを特徴づけることができます(図3)。解析の結果、野生型では茎の基部に応力が集中しており(図3B, C)、植物体全体を1点で支えるのに都合が良いことがわかりました。一方で、変異体では茎の中間部に引張応力と圧縮応力が不均一に分布する領域が生じることがわかり(図3E, F)、曲がり過ぎることによって、1点で支えられるような都合の良い形状から逸脱してしまうことが明らかになりました。この結果は、野生型の植物体でブレーキをかけてまっすぐな姿勢に戻る理由として、力学的に有利な姿勢であることを示しています。
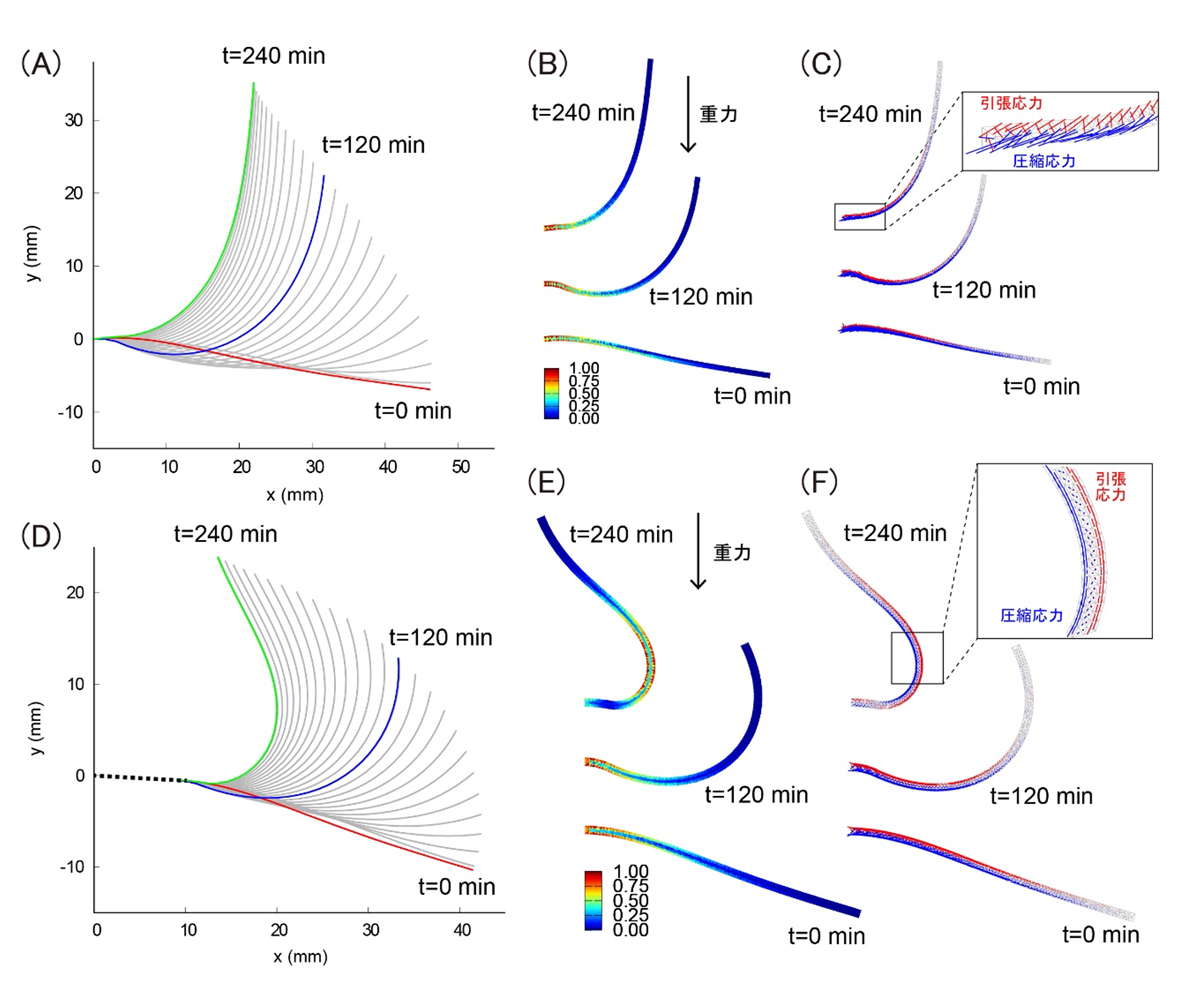 図3: 有限要素法による力学モデルの解析結果。
図3: 有限要素法による力学モデルの解析結果。
(A,D)野生型モデル(A)と変異体モデル(D)について0分、120分、240分で解析を行った。(B,E)植物体の自重によるフォンミーゼス応力分布。野生型では茎の基部に応力が集中し(B)、変異体では茎の中間部に応力が集中する(E)。(C,F)植物体の自重による応力方向の分布。野生型では茎の基部で上側に引張応力、下側に圧縮応力が生じるのに対し(C)、変異体では茎の中間部の右側に引張応力、左側に圧縮応力が生じる(F)。
・今後の期待
本研究により、植物生理学的なデータに基礎を置いた数理力学モデルを考察する方法論を確立できました。この実験データと数理モデルの連携によって、植物の形の変化だけからは推定できない、重力感受力パラメータを抽出することができ、さらにその抽出結果を内皮細胞におけるアミロプラスト運動のような生理学データによって再検証することを可能にしました。また、有限要素法による力学モデルによってどこに特異的な力が生じるかがわかるため、ダーウィンも注目した植物が姿勢を正す現象に対して、「力学的に有利であるから」といった新たな視点からの示唆を与えることができます。このような植物生理学データと力学モデルの連携によって、力学の視点から植物科学を考える新たな学術領域の創成へつながることが期待されます。
■ 用語解説
(1)重力屈性
植物を水平にしたときに,茎などの地上部は重力に抗して上に曲がり,地下部にある根は重力にしたがって下に曲がる現象を指す。古くは1806年にトーマス・ナイトが最初に報告し、1880年にダーウィンが情報伝達モデルを提案したことで知られている。
(2)自己受容性感覚
1907年にチャールズ・シェリントン(1932年ノーベル生理学賞受賞)が自己受容性感覚(固有覚)を「自分自身で刺激を与え自分自身でそれを受容する身体の感覚」と定義した。すなわち、体(またはその一部)の動き、姿勢、位置などを感じる能力を指す。近年、この言葉は、植物体が曲がり過ぎない姿勢を維持する姿勢制御の意味で多く用いられている。
(3)ミオシン
細胞骨格分子であるアクチン繊維の上を滑り運動するモータータンパク質である。植物細胞では、細胞内運動として有名な原形質流動を担う。動物細胞では,筋肉を構成する微小繊維として知られている。
(4)弾性棒理論
棒状構造が大きく変化する際に生じる力、ひずみ、変位を解析する力学理論を指し、主に材料力学や構造工学の分野で用いられる手法である。
(5)偏差成長
植物ホルモンのオーキシン(後述)の不均一分布によって生じる植物器官内の偏差的成長を指す。
(6)オーキシン
植物の成長を制御していると考えられている植物ホルモンのこと。
(7)有限要素法
主に時空間的な変動を予測する数値解析手法である。解析的に解くことが難しい方程式の近似解を数値的に得る方法として知られる。
■ 研究体制と支援
本研究は、津川暁(秋田県立大学,責任著者)、上田晴子(甲南大学,責任著者)、西村いくこ(京都大学,甲南大学)、出村拓(奈良先端科学技術大学院大学)、森田(寺尾)美代(基礎生物学研究所)、豊田正嗣(埼玉大学)の共同研究として行われました。
本研究は,文部科学省の科学研究費補助金 (JP18H05484, JP18H05489, JP18H05496, JP20K15832, JP19K06732, JP15H05776)、国立研究開発法人の戦略的創造研究推進事業(CREST(JPMJCR2121))、甲南学園平生太郎基金科学研究奨励助成金の支援を受けて行われました。
■ 問い合わせ先
<研究に関すること>
秋田県立大学システム科学技術学部機械工学科
助教 津川 暁(つがわ さとる)
<報道担当>
秋田県立大学広報・情報チーム
甲南学園広報部
基礎生物学研究所広報室
")
