
2018-01-19 国立研究開発法人量子科学技術研究開発機構
発表のポイント
・多能性幹細胞※1において、初めて糖鎖の一種のO-GlcNAc※2による分化※3の抑制機構を解明
・多能性幹細胞を再生医療に応用するために必要となる、未分化※3状態を安定的に維持して細胞を培養する方法の確立などへの応用が期待される
概要
国立研究開発法人量子科学技術研究開発機構(理事長 平野俊夫。以下「量研」という。)放射線医学総合研究所 放射線治療研究部 組織再生治療研究チームの三浦太一研究員と創価大学の西原祥子教授らは、マウス胚性幹細胞(ES細胞)※4を用いて多能性幹細胞の分化を促進するシグナルの活性化を「糖鎖」が抑制する新たなメカニズムを発見しました。
ES細胞は、様々な組織の細胞に分化できる能力(多能性)を有する、未分化性を維持した多能性幹細胞です。多能性幹細胞を再生医療に応用するためには、「未分化性を安定的に維持しつつ増やす」ことが必要です。この方法を樹立するためには、そもそも多能性幹細胞がどのように分化を抑制し未分化性を維持しているのか、そのメカニズムを解明することが必要であり、近年、盛んに研究が行われています。
ES細胞の未分化・分化の状態を決定する主要因子として「シグナル」が挙げられます。細胞にとってシグナルとはその名の通り「伝令」であり、シグナル構成因子として働く多種多様な細胞内の分子が次々に活性化することで最終的に細胞の状態を決定しています。ES細胞が未分化性を維持するためには「分化を促進するシグナル」が抑制されている必要がありますが、そのメカニズムについては不明な点が多くありました。
マウスES細胞の分化を促進するFGF4シグナルは、細胞内のシグナル構成因子がリン酸化されることで活性化します。つまり、FGF4シグナルにおいて、シグナル構成因子のリン酸化は活性化スイッチをONにするために必須なイベントであり、このリン酸化が抑制されると活性化スイッチはOFFになります。
研究グループは、細胞内のシグナル構成因子のリン酸化修飾と競合関係にある糖鎖修飾に着目し、研究を行ってきました。解析の結果、O-結合型N-アセチルグルコサミン(O-GlcNAc)の糖鎖修飾によるFGF4シグナル構成因子のリン酸化抑制が、マウスES細胞においてFGF4シグナルの活性化スイッチをOFFにしていることを発見しました。また、(1)O-GlcNAcを減少させるとFGF4シグナルが活性化すること、(2)FGF4シグナルの構成因子のうち、プロテインキナーゼC ζ(PKCζ)がO-GlcNAc修飾を受けてリン酸化が抑制されていること、(3) PKCζのリン酸化抑制がFGF4シグナルの活性化を抑制し、マウスES細胞の分化を抑制していることを初めて明らかにしました。
本研究は、多能性幹細胞において、初めてO-GlcNAcによる分化促進シグナルの制御機構を解明したものであり、これらの成果は、安定的に未分化性を維持できる多能性幹細胞の培養法の樹立につながることが期待されます。
本研究の一部は日本私立学校振興・共済事業団 学術研究振興資金、及び平成27年度 笹川科学研究助成の支援を受けました。
この成果は、米国科学雑誌「Stem Cell Reports」オンライン版に2017年12月15日に掲載されました。
補足説明
【研究開発の背景と目的】
多能性幹細胞は、(1)様々な組織に分化できる能力(多能性)、(2)自分と同じ娘細胞に分裂する能力(自己複製能)、(3)未分化の状態を維持することができる能力を併せ持つ細胞です。発生の初期段階である胚性3.5日目の胚から樹立されたマウス胚性幹細胞(ES細胞)は多能性幹細胞であり、胚の初期発生メカニズム解明のための研究、および再生医療への応用を目指した研究に広く用いられています。
多能性幹細胞を再生医療に応用するためには、「未分化性を安定的に維持しつつ増やす」ことが必須です。これを確立するためには、多能性幹細胞が分化を抑制して未分化性を維持するメカニズムを解明することが必要であり、未分化・分化の状態を決定する主要因子である「シグナル」についても近年、盛んに研究が行われています。
マウスES細胞において、未分化・分化状態の決定に関わるシグナルの一つに、繊維芽細胞増殖因子-4(fibroblast growth factor 4: FGF4)シグナルがあります。FGF4シグナルは、分化の開始に必須なシグナルで、FGF4(リガンド)がFGF受容体に結合すると、それを合図に、次々と細胞内シグナル構成因子として働く分子がリン酸化されていきます。主要なFGF4シグナル構成因子にMEK(mitogen-activated protein kinase/extracellular signal-related kinase)、ERK1/2(extracellular signal-related kinase 1/2)があり、MEKは上流因子によりリン酸化されることで活性化し、リン酸化(活性化)MEKは下流のERK1/2をリン酸化します。リン酸化(活性化)ERK1/2はマウスES細胞の分化を促進するように働きかけます(図1)。しかし、マウスES細胞においてFGF4シグナルが抑制されているメカニズムについては不明な点が多くありました。

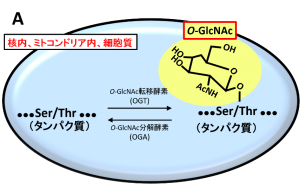
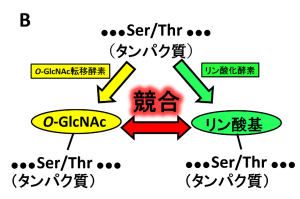
O-GlcNAc修飾は核内やミトコンドリア内、細胞質で行われる唯一の糖鎖修飾であり、O-GlcNAc転移酵素(OGT)は、細胞骨格タンパク、転写因子、シグナル構成因子などの種々のタンパク質のセリン(Ser)・スレオニン(Thr)残基にGlcNAcを一つ付加します(図2A)。セリン・スレオニン残基はリン酸化修飾を受ける部位でもあるため、O-GlcNAc修飾とリン酸化修飾は競合関係にあることから、リン酸化抑制がO-GlcNAc修飾の代表的な機能の一つとされています(図2B)。
マウスES細胞においてO-GlcNAcは未分化性に必須であること、分化に伴いO-GlcNAcが減少することなどが報告されていますが、まだ解明されていないことが多く、シグナルとの関連についても不明でした。そこで本研究は、マウスES細胞におけるO-GlcNAc糖鎖修飾によるFGF4シグナル抑制機構の解明を目的として研究を行いました。
【研究の手法と成果】
(1)O-GlcNAcのFGF4シグナルへの関与を探る
始めに、マウスES細胞でRNA干渉法※5を用いて、O-GlcNAc転移酵素(Ogt)をノックダウン(KD)し、O-GlcNAcの量を減少させました。Ogt KD細胞において、未分化性維持に必須な転写遺伝子群(OCT4、SOX2、NANOG)が減少し、分化が促進したことから、O-GlcNAcはマウスES細胞の未分化性維持に必須であることが分かりました(図3①)。
次に、O-GlcNAcのFGF4シグナルへの関与を探るために、Ogt KD細胞において、FGF4シグナルのうち、どの構成因子が活性化しているか検討しました。その結果、Ogt KD細胞でMEKとERK1/2のリン酸化が有意に増加していることが分かりました(図3②)。Ogt KD細胞にMEK阻害剤を添加してMEKの活性化を阻害すると、MEK、およびERK1/2のリン酸化が阻害され、未分化性維持に必須な転写遺伝子群が増加して分化が抑制されたことから、MEK-ERK1/2経路の活性化により分化が促進したことがわかりました(図3③)。また、通常のマウスES細胞と、Ogt KD細胞それぞれにFGF4を添加し、その刺激に対するMEKとERK1/2のリン酸化の促進を検討しました。その結果、Ogt KD細胞において、MEKとERK1/2のリン酸化が顕著に促進していることが分かりました(図3④)。
以上より、マウスES細胞において、O-GlcNAcが減少すると、FGF4シグナルが活性化し、分化が促進し、結果的に未分化性が失われることが分かりました。
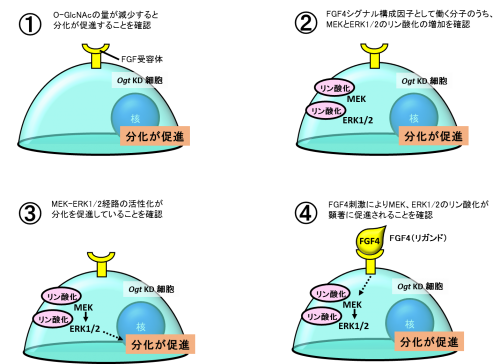
図3 Ogt KD細胞を用いたO-GlcNAcのFGF4シグナル経路への関与を解析した実験
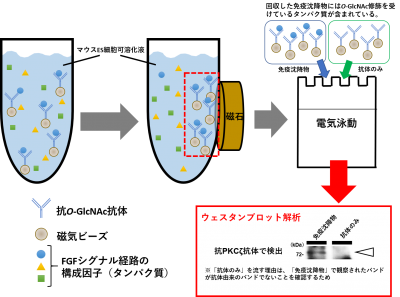
図4:抗O-GlcNAc抗体を用いた免疫沈降解析
最後に、PKCζのどの部位にO-GlcNAcが結合しているのかを検討しました。PKCζは410番目のスレオニン(T410)がリン酸化されることで活性化します。そこで、このT410がO-GlcNAc修飾されることでリン酸化が抑制されているのではないかと考え、Flagタグのついた野生型のPKCζと、同じくFlagタグのついたT410をアラニンに置換したPKCζ(PKCζ T410A)をそれぞれマウスES細胞で発現させた後に抗Flag抗体を用いて免疫沈降物を収集し、抗O-GlcNAc抗体を用いてウェスタンブロット解析を行いました。PKCζ(野生型)の免疫沈降物を電気泳動し、抗O-GlcNAc抗体を用いて検出したところ、PKCζの分子量の位置(図5の矢頭上)にO-GlcNAcのバンドを確認することができ、PKCζ(野生型)はO-GlcNAc修飾を受けていることが改めて明らかとなりました。また、免疫沈降されたPKCζ(図5の矢頭下)に対するPKCζ上のO-GlcNAc(図5の矢頭上)のバンドの濃さを、PKCζ(野生型)とPKCζ T410Aの免疫沈降物とで比べたところ、PKCζ T410AではPKCζ(野生型)よりも、O-GlcNAcのバンドが薄いことが分かりました。これは、410番目のスレオニンをアラニンに置換したことでO-GlcNAc修飾がされなくなったことを意味し、このことから、PKCζのT410がO-GlcNAc修飾を受ける部位であることがわかりました(図5)。
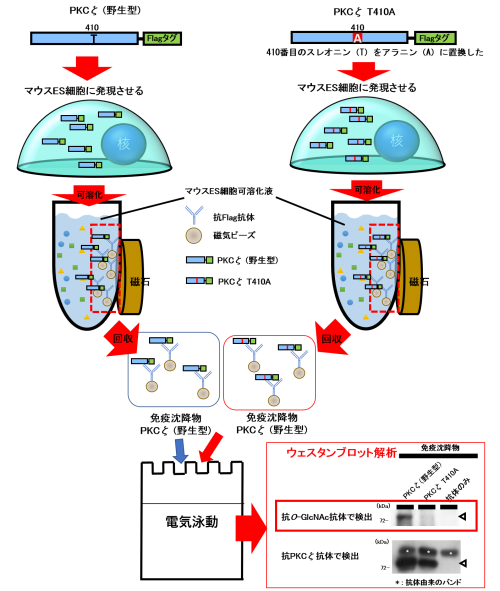
図5:免疫沈降解析によるPKCζのO-GlcNAc修飾部位の同定
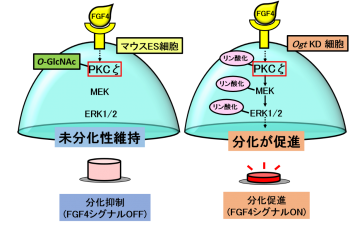
図6:O-GlcNAcによるFGF4シグナル抑制機構
今後の展開
本研究により、多能性幹細胞において、初めてO-GlcNAcによる分化促進シグナルの制御機構が明らかとなりました。今後は、マウスES細胞において未分化性維持や分化促進に関わるFGF4以外のシグナルにおいてもO-GlcNAcが制御しているか検討する予定です。マウスES細胞でO-GlcNAcによる分化・未分化シグナルの制御機構をより詳細に明らかにすることは、ヒトの多能性幹細胞におけるO-GlcNAcを含めた糖鎖修飾と未分化性の維持の関連を解明する重要な足がかりとなり、未分化性を安定的に維持できる多能性幹細胞の培養法や、効率的な分化法の確立、そして、多能性幹細胞を用いた再生医療の一つとして、放射線障害治療にも応用できることが期待されます。
【用語解説】
※1量子科学技術研究開発機構(理事長 平野俊夫。以下「量研」という。)放射線医学総合研究所 放射線治療研究部 組織再生治療研究チームの三浦太一研究員と創価大学の西原祥子教授らは、マウス胚性幹細胞(ES細胞)※4を用いて多能性幹細胞の分化を促進するシグナルの活性化を「糖鎖」が抑制する新たなメカニズムを発見しました。
多能性幹細胞は、(1)様々な組織に分化できる能力(多能性)、(2)自分と同じ娘細胞に分裂する能力(自己複製能)、(3)未分化の状態を維持することができる能力を併せ持つ細胞です。胚性幹細胞(ES細胞)や人工多能性幹細胞(iPS細胞)は多能性幹細胞です。
※2 O-結合型N-アセチルグルコサミン(O-GlcNAc)
O-結合型N-アセチルグルコサミン(O-GlcNAc)修飾は核内やミトコンドリア内、細胞質で行われる唯一の糖鎖修飾であり、O-GlcNAc転移酵素(OGT)は、細胞骨格タンパク、転写因子、シグナル構成因子などの種々のタンパク質のセリン(Ser)・スレオニン(Thr)残基にN-アセチルグルコサミン(GlcNAc)を一つ付加します。O-GlcNAcはO-GlcNAc分解酵素(OGA)により分解されます。O-GlcNAcの機能についてはまだ不明な点が多くありますが、代表的な機能としてリン酸化抑制が挙げられます。セリン・スレオニン残基はリン酸化修飾を受ける部位でもあるため、O-GlcNAc修飾とリン酸化修飾は競合関係にあることから、O-GlcNAcがリン酸化を抑制していると考えられています。
※3 分化・未分化
多能性幹細胞において、「分化する」ということは、「ある組織の細胞になる」ということです。例えば、「ES細胞が神経細胞に分化する」や、「iPS細胞が筋肉の細胞に分化する」という表現を使います。一方、多能性幹細胞は、どの組織の細胞にも分化していない状態、言い換えれば、どの組織の細胞に分化するかまだ決定していない状態であり、この状態を「未分化」な状態と言います。
※4 胚性幹細胞(ES細胞)
胚性幹細胞(ES細胞)は、発生初期段階の胚の中にある内部細胞塊から樹立された多能性幹細胞です。内部細胞塊は将来、私たちの体を構成するすべての組織の細胞に分化できる能力(多能性)を持っているため、ES細胞も同様の能力を持っています。
※5 RNA干渉法
RNA干渉とは、2本鎖RNAと相補的な塩基配列を有するメッセンジャーRNA(mRNA)が分解される現象のことです。このRNA干渉を利用することで、人為的にある特定の遺伝子の量を減少させる(ノックダウンする)ことできます。
※6 抗O-GlcNAc抗体
O-GlcNAc修飾を認識し、結合する抗体です。O-GlcNAc研究において必ず使用される最重要のツールです。
※7 免疫沈降解析
細胞可溶化液などに、あるタンパク質を特異的に認識する抗体を添加し、その抗体が結合したタンパク質のみを精製する方法です。カラムを用いる方法や磁気ビーズを用いる方法があります。
※8 ウェスタンブロット解析
細胞可溶化液などのタンパク質の混合液から、特定のタンパク質のみを検出する方法です。検出には、主に、特定のタンパク質に対する抗体(一次抗体)と、その一次抗体に結合する酵素標識のされた二次抗体を用います。特定のタンパク質は、そのタンパク質の分子量の位置にバンドとして検出することができます。バンドの濃淡から特定のタンパク質の発現量や、リン酸化されたタンパク質に対する抗体を一次抗体として用いれば、特定のタンパク質のリン酸化状態を検討することができます。

