
2019-10-16 東京大学
河野 優(生物科学専攻 特任助教)
川口 ひかる(神奈川大学大学院理学研究科生物科学専攻 学生)
水澤 直樹(法政大学生命科学部生命機能学科 教授/法政大学マイクロナノテクノロジー研究センター 兼担研究員)
矢守 航(生物科学専攻 准教授)
鈴木 祥弘(神奈川大学大学院理学研究科生物科学専攻 准教授)
寺島 一郎(生物科学専攻 教授)
発表のポイント
- 遠赤色光(注1)は、単独では光合成を駆動しないため、その光合成への影響は研究されてこなかった。本研究は、光合成を駆動できる波長域の光の強さ(注2)が頻繁に変化する変動光環境下において、遠赤色光が光合成の効率(注3)を上昇させることを明らかにした。
- 多くの研究において植物に照射される光合成駆動光の強度は一定であることが多い。このような定常光条件下では、遠赤色光は光合成にほとんど影響を与えない。本研究は、光合成を直接には駆動しない遠赤色光が、植物の光合成能を最大限に引き出すことを見出した。
- 遠赤色光による光合成増強(促進)機構の解明は、将来予想される食料不足問題解決に向けた光合成能増大作物の創出に新たな可能性を与えることが期待される。また、植物工場などにおける“光照射法”の最適化にも大きく貢献する。
発表概要
陸上植物は波長400 – 700 nmの青色〜赤色の領域の光、つまり可視光領域の光を吸収して光合成を行うので、この波長域の光は光合成有効放射とよばれる。一方、人の目には見えないとされる遠赤色光とよばれる700 – 800 nmの領域の光は、植物の発生や形態形成に大きく作用するが、光合成との関連ではその作用はほとんど研究されてこなかった。確かに、700 nmより長波長の単色光はほとんど光合成を駆動しない。一方、太陽光は遠赤色光を豊富に含む。また、植物の浴びる太陽光の強度は、雲による遮蔽や、上部に存在する植生によって、頻繁かつダイナミックに変化する。東京大学大学院理学系研究科の河野優特任助教らは、アブラナ科のモデル植物シロイヌナズナの葉に、野外変動光を模した光合成駆動光に加えて遠赤色光を照射したときの光合成応答を解析した。その結果、遠赤色光の補光によって光合成速度が増加すること、遠赤色光の効果は光の強さが頻繁に変化する変動光環境下で顕著に見られることが明らかになった。単独では光合成を駆動しない遠赤色光は、光合成の調節に深く関わっており、光エネルギーや電子の渋滞緩和のための交通整理の役割を果たしているのである。今回の結果は、食料不足問題の解決に向けた光合成能増大作物の創出に新たな可能性を与えるものである。
発表内容
光合成に利用される光の範囲は400 – 700 nmの波長域に限られており、光合成有効放射とよばれる。光によって、2つの光化学系(IIとI)が励起され、光合成が進行する(図1B)。一方、700 – 800 nmの波長域の光は遠赤色光とよばれる。700 nmより長波長の単色光は、光合成によるO2発生やCO2固定を駆動しないが、光化学系Iを励起することはできる。この意義は不明だが、光合成によるO2発生やCO2固定を駆動しないためか、光合成における遠赤色光の役割は長年無視されてきた。地上に降り注ぐ太陽光には、光合成有効放射だけでなく遠赤色光も豊富に含まれている。さらに、野外光環境は、光の強さ(光強度)と質(波長特性)が遮蔽物の影響によって絶えず変化する“変動光環境”である(図1A)。植物は、光を吸収して光合成をする一方で、光の過剰吸収で光合成装置(特に光化学系)が壊れないようにするために光エネルギーを熱として散逸する機構(光化学系II活性の下方制御;注4)を有している。変動光に曝される野外植物は、吸収した光の光合成利用と熱散逸とを状況に応じて使いわけていると考えられる。しかし、これまでの光合成測定は、遠赤色光を含まない光合成有効放射を照射して行われてきた。また、光強度が変化しない定常光下で行われるのが一般的であった。このような条件下で得られた結果が野外の植物の真の光合成応答を反映しているのかは疑問である。本研究を中心になって進めてきた東京大学理学系研究科の河野優特任助教、寺島一郎教授らは、変動光は植物にとってストレス要因になりうることをすでに見出しており、変動光下で効率よく光合成を行うことは植物にとって重要な戦略であると考えた。遠赤色光が光化学系Iにのみ作用することから、変動光下の光合成応答における遠赤色光の役割を精査すべく本研究を行った。
モデル植物のアブラナ科シロイヌナズナを、光や温度、湿度等が制御された植物育成室で栽培し、供試した。光合成励起光と遠赤色光の光源には発光ダイオードを用いた。変動光はプログラムで厳密に制御して照射した(図1C)。光合成を測定するための専用の装置(同化箱)内におかれた葉の光合成によるCO2濃度減少と蒸散による水分濃度増加を測定し、葉のCO2交換速度を見積もった。さらに、光合成反応の初期反応の場である光化学系IIの活性と光化学系IIの熱散逸機構の活性(NPQ)とを、パルス変調クロロフィル蛍光測定法(注5)によって算出した。


図1:(A)野外晴れの日における光強度の日変化。(B)光合成有効放射と遠赤色光が光合成反応に与える影響を示した模式図。光合成有効放射は光化学系IIとIを励起することによって、光合成を駆動する。一方、遠赤色光は光化学系Iのみを励起する。強光下、強すぎる光の一部は、熱に変換されて安全に散逸される。弱光下、ほとんどの吸収した光エネルギーは光合成に利用される。(C)実験に用いたモデル植物のアブラナ科シロイヌナズナ。
強光と弱光を繰り返す変動光下の光合成速度(図2A)と光化学系IIの活性(図2B)およびNPQ(図2C)は、強光で高くなり、弱光では低下する。いくつかの酵素反応からなる光化学系IIの熱散逸機構は弱光下の光合成を抑制してしまうので、速やかに解消されなければならない。遠赤色光補光の効果(光合成速度増加)は、変動光の強光期では見られなかった。一方で、強光期から弱光期に切り替わると、光合成速度と光化学系IIの活性は、遠赤色光補光により有意に増加した。NPQの解消も遠赤色光存在下で速かった(図2)。遠赤色光の補光効果を光強度一定の定常光下で解析したところ、光合成とNPQの変化はわずかであった。これらの結果から、遠赤色光による光合成速度促進効果は、吸収した光エネルギーの利用(光合成)と廃棄(熱散逸)の素早い切替によって達成されること、変動光下で強光から弱光に切り替わった際に顕著であることが明らかになった。
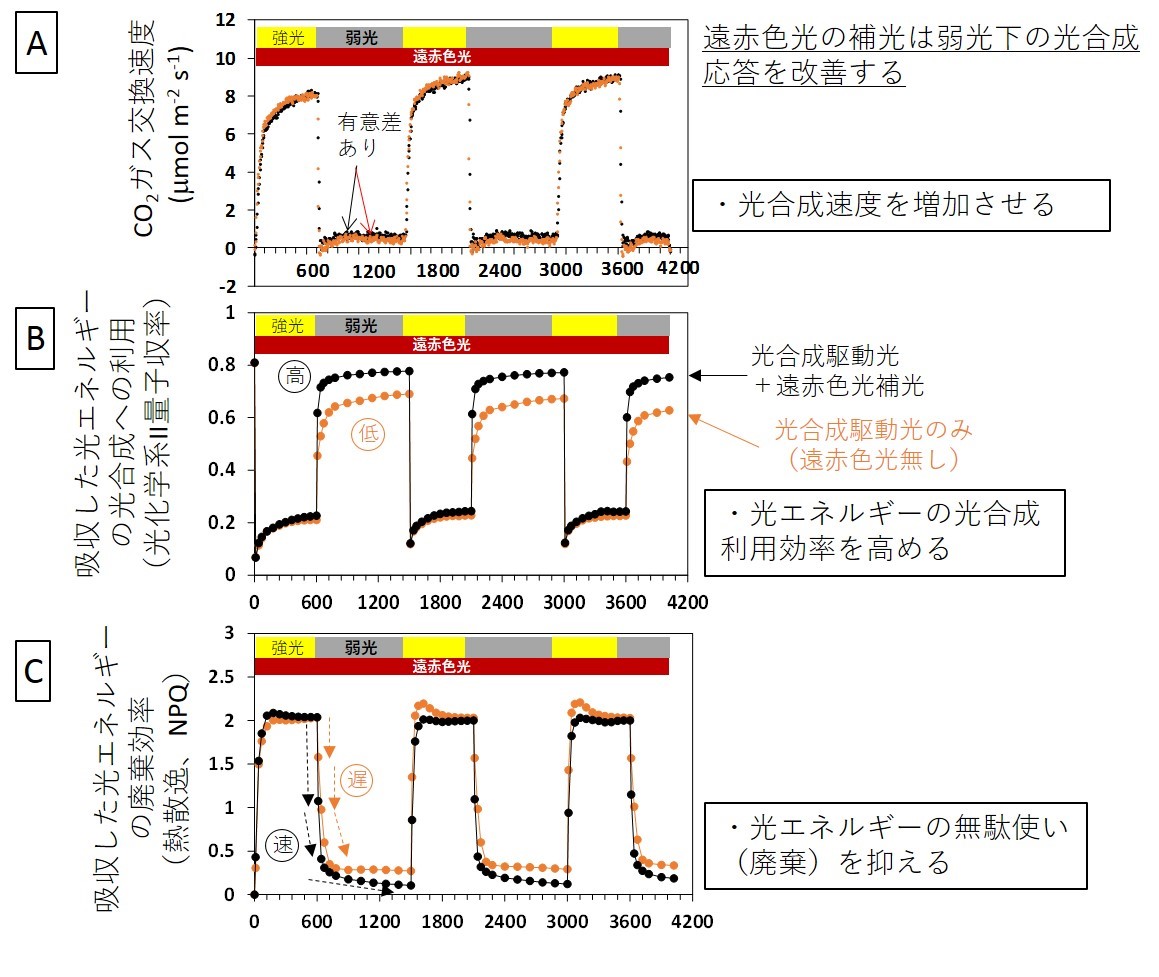
変動光照射時間(秒)
図2:変動光中の光合成応答を遠赤色光の補光あり(黒色)と補光なし(橙色)で比較した。
変動光中に遠赤色光が存在すると、強光から弱光に切り替わったときのCO2ガス交換速度が高い(A)。これは、遠赤色光によって、吸収した光エネルギーの光合成利用(B)と廃棄(C)の速やかな切り替えが行われたためである。
光合成を駆動しないとされてきた遠赤色光が、変動光下の光合成の調節に有効であることを明らかにした本研究成果は、光合成の研究、すなわち、植物光合成応答の真の理解には遠赤色光を考慮することが必須であることを意味している。今後は、遠赤色光による光合成促進機構の徹底解明を目指す。本研究成果と光合成促進機構の解明は、植物を扱う上での“光照射法”に再考を促すだけでなく、将来の食糧不足問題解決に向けた光合成能強化作物の創出へ貢献することが期待される。
本研究は、科学研究費補助金である若手研究、および新学術領域研究「新光合成:光エネルギー変換システムの再最適化」のサポートを受けて行われました。
発表雑誌
- 雑誌名
Plant and Cell Physiology論文タイトル
Far-red light accelerates photosynthesis in the low-light phases of fluctuating light著者
Masaru Kono*, Hikaru Kawaguchi, Naoki Mizusawa, Wataru Yamori, Yoshihiro Suzuki and Ichiro Terashima*DOI番号
10.1093/pcp/pcz191

用語解説
注1 遠赤色光
赤外線の一部で、700 – 800 nmの波長域の光のこと。おもに植物を扱う研究において用いられる用語である。植物の伸長成長や発芽制御などには遠赤色光が関わるため、その効果はよく調べられている。遠赤色光はクロロフィルにも吸収されるが、光合成有効放射に比べて、その吸収率は小さい。また、光化学系IIのクロロフィルにはほとんど吸収されないため、遠赤色光単独では光合成による O2発生やCO2固定をほとんど駆動しない。
注2 光の強さ
光合成反応では、光を粒子(光量子)としてみたときに、光量子がどの程度のエネルギーを持っているのかではなく、光量子の数が重要となる。そのため、光合成分野では、波長400-700 nmの光量子数を基本とした単位、光量子束密度(μmol photons m-2 s-1)を使う。一般的に、室内の蛍光灯の下に立った時が10 μmol photons m-2 s-1程度、深い森の中だと10 – 50 μmol photons m-2 s-1程度、曇り空だと50 – 400 mmol photons m-2 s-1、真夏の直射日光がおよそ2000 μmol photons m-2 s-1程度になる。
注3 光合成の効率
光合成速度は葉に照射される光の強さに比例して高くなる。しかし、ある程度の強度に達すると光合成速度は飽和する。光合成を飽和させるレベルよりも強い光が照射されると、過剰な光エネルギーが発生する。植物は、過剰な光エネルギーを熱に変換して安全に散逸機構を持っている。強すぎる光のもとでは、吸収した光エネルギーを率先して熱散逸してやることで、自身の光合成装置が壊れないようにしている。
注4 光化学系IIの下方制御
光合成に利用できる以上の光エネルギーを光化学系IIに届けないようにするための防御応答。過剰な光エネルギーを熱に変換して散逸することで、光化学系IIの活性を下方制御(ダウンレギュレート)する。光化学系IIでの反応を光化学反応というのに対し、下方制御は光を光合成以外に使う(熱にしてしまう)という意味で非光化学的消光(non-photochemical quenching略してNPQという)ともいう。NPQはいくつかの酵素反応が関与するため、強光下での誘導と弱光後の解消には時間的遅延が生じる。強光から弱光に切り替わった直後に、NPQが速やかに解消しないと、光合成に利用できる弱光のエネルギーまでも熱として逃がしてしまうため、光合成速度の低下につながる。
注5 パルス変調クロロフィル蛍光測定法
葉に吸収され光化学系IIに流れ込む光エネルギーは、光合成に使われるか、熱となって散逸するか、クロロフィル蛍光として再放出される。したがってクロロフィル蛍光の収率は、光合成や熱散逸の効率に依存して変化する。パルス変調された測定光に対応して放出された測定光と同調したクロロフィル蛍光だけをモニターすることで、吸収した光エネルギーの光合成もしくは熱散逸に流れる割合を相対評価できる方法がパルス変調蛍光法である。この方法を使うことで、非破壊的に光合成活性を評価できる。
―東京大学大学院理学系研究科・理学部 広報室―

