遠隔記憶は大脳皮質と海馬の相互作用により想起される
2019-12-18 理化学研究所
理化学研究所(理研)脳神経科学研究センター神経回路・行動生理学研究チームのトーマス・マックヒューチームリーダー、牧野祐一基礎科学特別研究員(研究当時)らの研究チームは、形成されてから長い時間が経った記憶(遠隔記憶[1])の想起時には大脳皮質[2]と海馬[3]の間に多様な電気生理的相互作用が起こり、大脳皮質が海馬に残存する記憶痕跡[4]の活性化を手助けすることで、記憶の想起が可能になっていることを発見しました。
本研究成果は、記憶形成からの時間経過により複雑に変化する記憶の脳内メカニズムの全容解明につながると期待できます。
海馬で形成された記憶は、年月を経て遠隔記憶になるにつれて次第に大脳皮質に移動することが知られています。しかしその結果、海馬が記憶の想起に不要になるのかどうかははっきりと分かっていません。
今回、研究チームはマウスが遠隔記憶を想起している間の、大脳皮質と海馬における神経活動を同時に記録しました。その結果、記憶想起中に両領域の活動が複数の周波数帯で同期し、海馬の一部の神経細胞の活動を促進していることが明らかになりました。このことは、時間が経っても海馬には記憶の痕跡がまばらに残っており、大脳皮質がその活性化を手助けすることで記憶想起を促すことを示唆しています。さらに、この同期信号を機械学習アルゴリズム[5]で解析したところ、記憶の古さを判定できることが分かり、記憶形成からの経過時間を客観的に示すバイオマーカーとして利用できる可能性が示されました。
本研究は、米国の科学雑誌『Cell Reports』(12月17日付:日本時間12月18日)に掲載されます。

図 遠隔記憶想起時の大脳皮質と海馬の相互作用
背景
脳は、さまざまな記憶を数カ月、数年という長期間保つことができますが、ある種の記憶は形成されたときから同じ脳部位に保存され続けるわけではなく、時間が経ち、遠隔記憶になるにつれて、次第に異なる脳部位に移動することが知られています。特に過去の体験についての記憶(エピソード記憶)は、形成された直後(近時記憶[1])は海馬に保存されるものの、時間経過とともに大脳皮質に移動し、そこで遠隔記憶として固定されると考えられてきました。この考え方は「記憶の標準固定化説[6]」と呼ばれ、数十年にわたって支持されてきました。
一方で、エピソード記憶は、遠隔記憶になっても全てが大脳皮質に移動するのではなく、部分的に海馬に残り続けるという研究報告も多数あります注1)。これらの報告から、遠隔記憶の痕跡は大脳皮質と海馬を含む複数の脳部位で保持されるという「記憶の多重痕跡説[6]」が提唱されています。しかし、遠隔記憶の想起時に大脳皮質と海馬がそれぞれどのような役割を果たすのか、また両部位の間で具体的にどのような情報伝達が行われて遠隔記憶が想起されるのかは分かっていませんでした。
この課題に取り組むため、研究チームはマウスが遠隔記憶を想起している間の大脳皮質と海馬の活動を同時に記録し、両部位間での電気生理的相互作用を詳細に調べました。
注1)Winocur, G., Moscovitch, M., and Bontempi, B. (2010). Memory formation and long-term retention in humans and animals: convergence towards a transformation account of hippocampal-neocortical interactions. Neuropsychologia. 48, 2339-2356.
研究手法と成果
まず研究チームは、複数の脳部位から神経活動を記録できる多電極アレイを、マウスの大脳皮質の一部である前帯状皮質[2]と海馬CA1領域[3]に埋め込みました(図1A, B)。そして、恐怖条件付け[7]を用いてマウスに恐怖記憶を形成させ、その1日後(近時記憶)と1カ月後(遠隔記憶)にマウスが恐怖記憶を想起しているときの神経活動を両部位で記録しました(図1C)。
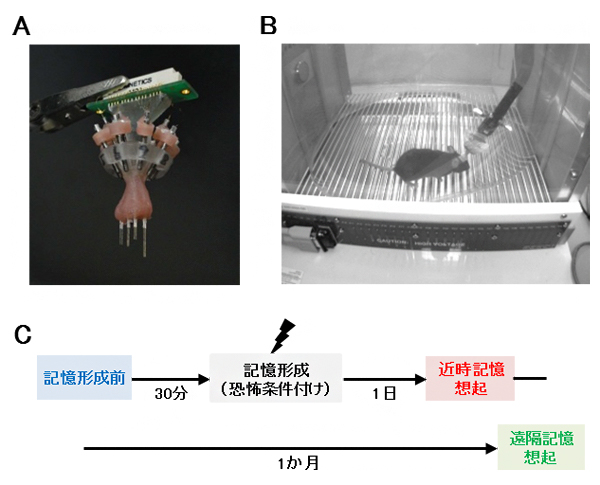
図1 記憶想起時の脳活動の多領域同時記録
A.脳の多領域の活動を同時に記録するための多電極アレイ。8本の電極を複数の領域に挿入し、それぞれを上下方向に動かすことによって、単一の神経細胞と細胞集団の活動を検出する。
B.チャンバーの中で形成された恐怖記憶を想起しているマウス。脳に埋め込まれた電極からの活動を、接続されたケーブルを通して記録する。
C.実験全体の流れ。まず記憶形成前の脳活動を記録し、30分後にチャンバー内で恐怖条件付けを行うことにより恐怖記憶を形成する。その1日後(近時記憶)と1カ月後(遠隔記憶)に、チャンバーで恐怖記憶を想起しているときの脳活動を再び記録する。
多くの脳部位では、アルファ波などの例で知られるように、神経細胞の集団が特定の周波数で周期的に活動することが分かっています。そこで、記憶想起時の前帯状皮質と海馬CA1領域での周期的活動を測定したところ(図2A)、前帯状皮質と海馬CA1領域それぞれでシータ波[8](6~12Hz)の活動が見られ、かつそれが両部位間で同期していることが分かりました。そしてこの両部位間での同期は、記憶形成前や近時記憶の想起時に比べて、遠隔記憶の想起時に高くなっていました(図2B)。さらに高ガンマ波[8](60~90Hz)においても同様に、両部位間の同期が遠隔記憶想起時に強くなることが分かりました(図2C)。これらの結果から、遠隔記憶の想起時には前帯状皮質と海馬CA1領域が複数の周波数帯で同期し、想起を促している可能性が示されました。
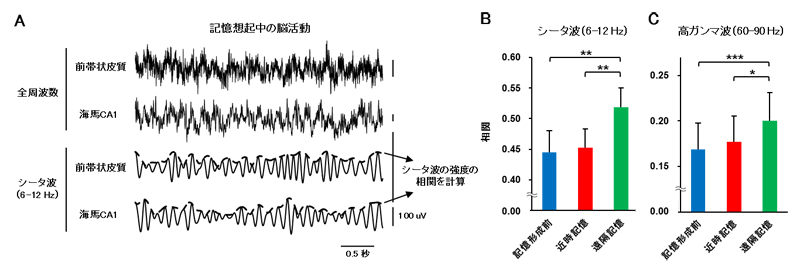
図2 遠隔記憶想起時の前帯状皮質と海馬CA1領域の活動同期
A.記憶を想起しているときの、前帯状皮質と海馬CA1領域の神経細胞集団の活動の例。上の二つのトレースは生の記録データ(全周波数)、下の二つはシータ波を取り出したもの。Bでは、このシータ波の強度の部位間での相関を計算している。
B.記憶形成前または想起時の、前帯状皮質と海馬CA1領域の間でのシータ波強度の相関。遠隔記憶時に相関が強くなった。
C.記憶形成前または想起時の、前帯状皮質と海馬CA1領域の間での高ガンマ波強度の相関。遠隔記憶時に相関が強くなった。
次に、このような脳部位間での相互作用が、どのようにして記憶痕跡に働きかけているのかを知るために、エピソード記憶において記憶痕跡を保持していると考えられている個々の神経細胞のスパイク発火[9]を詳細に調べました。その結果、海馬CA1領域のうち一定割合の神経細胞は、遠隔記憶想起時に前帯状皮質の活動と同期して発火することが分かりました(図3A)。また、前帯状皮質の活動と同期して発火する神経細胞同士は海馬CA1領域の中で離れて存在し、遠隔記憶想起時に同時に発火していました(図3B)。これらの結果から、遠隔記憶において海馬は少数の神経細胞で記憶痕跡を保持しており、前帯状皮質はそれらの細胞の発火を制御する、つまり記憶痕跡の活性化を手助けすることで、記憶を呼び起こしている可能性が示されました。
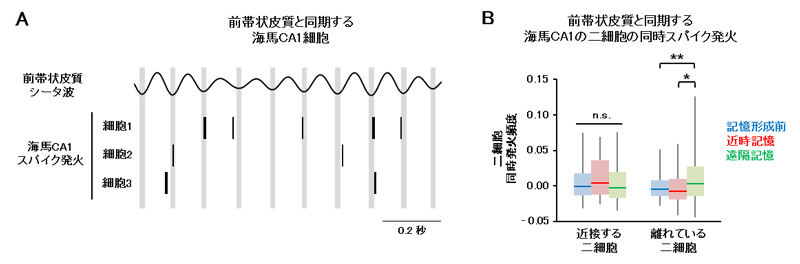
図3 遠隔記憶想起時の海馬二細胞の同時スパイク発火
A.海馬CA1領域における個々の神経細胞のスパイク発火の例。図に示されている三つの神経細胞は、前帯状皮質におけるシータ波の特定のフェーズ(縦線で示されている谷の部分)と同期して発火している。
B.海馬CA1領域における二つの神経細胞の同時スパイク発火。CA1領域の中で近接する二細胞の間では同時発火が見られないものの、300マイクロメートル(1マイクロメートルは1,000分の1ミリメートル)程度離れている二細胞の間では、遠隔記憶想起時に同時発火が見られる。
以上から、遠隔記憶の想起時には、前帯状皮質と海馬CA1領域の間に、近時記憶想起時とは異なる電気生理的相互作用が起こることが分かりました。また、この結果は両部位間での相互作用が記憶の古さを示すバイオマーカーとなることを示唆しています。
この可能性を模索するため、記憶想起時の前帯状皮質と海馬CA1領域の間での同期信号を周波数ごとに分け、機械学習アルゴリズムに入力して解析を試みました(図4A)。その結果、シータ波・低ガンマ波[8]・高ガンマ波の三つの同期信号の組み合わせから、想起していた記憶が近時記憶と遠隔記憶のどちらだったかを正確に判定することに成功しました(図4B)。これにより、記憶が形成された時期が分からなくても、それを想起しているときの脳活動を測定することで形成時期(記憶の古さ)を推定するという全く新しい手法が見いだされました。
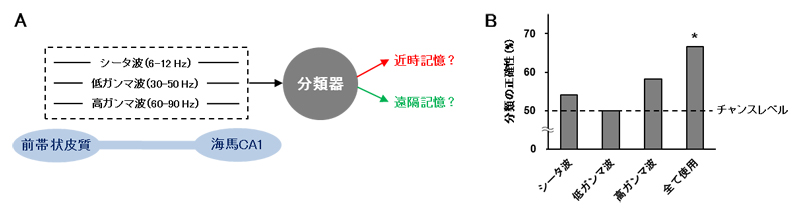
図4 機械学習による近時記憶と遠隔記憶の分類
A.記憶想起時の前帯状皮質と海馬CA1領域の間での同期信号から、近時記憶と遠隔記憶を区別する機械学習の模式図。
B.各種の入力信号における近時記憶・遠隔記憶の分類の正確性。三つの信号を全て入力した場合に、偶然の結果(チャンスレベル)よりも統計的に有意に高い正確性が得られた。
今後の期待
本成果は、遠隔記憶の想起時には大脳皮質と海馬の間に多様な情報伝達があることを明らかにし、遠隔記憶の多重痕跡説を支持する重要な知見です。
今後、それぞれの脳部位で記憶のどのような側面が保存され、それらがどのように組み合わさって記憶が想起されるのかなどの詳細を明らかにすることにより、記憶想起メカニズムの包括的理解につながると期待できます。
補足説明
1.遠隔記憶、近時記憶
形成されてから間もない記憶を近時記憶、形成から長い時間が経った記憶を遠隔記憶という。明確な定義はないが、マウスやラットでは形成から1~2週間以内、ヒトでは数カ月以内の記憶を近時記憶、それ以降の記憶を遠隔記憶と区別することが多い。近時記憶と遠隔記憶は、関わる脳部位や維持・想起のメカニズムが異なることが広く知られている。
2.大脳皮質、前帯状皮質
大脳皮質は脳の最も外側の広範囲にわたる領域であり、知覚、記憶、思考などさまざまな機能に関わる。その一部分である前帯状皮質は前頭部に位置し、マウスやラットにおいて遠隔記憶に必要なことが分かっている。
3.海馬、CA1領域
内側側頭葉に位置する脳部位の海馬は、記憶や学習に深く関わる。特に海馬の一部分であるCA1領域は、過去の体験についての記憶(エピソード記憶)や空間情報の記憶(空間記憶)に関係することが知られている。
4.記憶痕跡
記憶の形成に伴い脳内に作られる、神経細胞の結合や構造的変化などの物理的な痕跡。この痕跡が脳内に残り続けることにより記憶が保たれ、また活性化されることにより記憶が想起されると考えられている。
5.機械学習アルゴリズム
サンプルデータをコンピュータに入力し、データに共通する法則やルールを抽出することにより、新しいデータについて予測・決定を行う方法。
6.記憶の標準固定化説、記憶の多重痕跡説
ある種の記憶(特にエピソード記憶)は、形成されてから時間が経ち、固定化されるにつれて海馬から大脳皮質に移動するという考え方を、記憶の標準固定化説と呼ぶ。一方、記憶は固定化された後でも、その異なる側面が海馬と大脳皮質の両方にそれぞれ保たれているという考え方を、記憶の多重痕跡説と呼ぶ。
7.恐怖条件付け
マウスに音や光などの手掛かりを与えているときに電気ショックを施し、与えた手掛かりに対し恐怖を感じる記憶を形成させる手法。手掛かりに対するすくみ反応(体を動かさずにしばらくじっとしている行動)を観察することにより、記憶想起の度合いを測定する。マウスやラットでの記憶研究に広く用いられている。特に、本研究でも用いられている状況(文脈)に恐怖を感じる状況恐怖記憶は、近時記憶から遠隔記憶になると海馬CA1領域から前帯状皮質に移動することが知られている。
8.シータ波、高ガンマ波、低ガンマ波
多くの脳部位における神経細胞集団の代表的な周期的活動。それぞれ学習、記憶、認知、感情などさまざまな機能に関わることが知られている。
9.スパイク発火
1ミリ秒以下の速さで起こる、神経細胞の最も基本的な電気的活動。この活動の頻度やタイミングが脳のさまざまな情報処理の基盤となっている。
研究チーム
理化学研究所 脳神経科学研究センター
神経回路・行動生理学研究チーム
チームリーダー トーマス・マックヒュー(Thomas J.McHugh)
基礎科学特別研究員(研究当時) 牧野 祐一(まきの ゆういち)
テクニカルスタッフⅠ デニス・ポリガロフ(Denis Polygalov)
視覚意思決定研究チーム
チームリーダー アンドレア・ベヌッチ(Andrea Benucci)
大学院生リサーチ・アソシエイト フェデリコ・ボラノス(Federico Bolanos)
研究支援
本研究の一部は、日本学術振興会(JSPS)科学研究費補助金基盤研究(S)「Elucidating the Dynamics of Memory(研究代表者:McHugh Thomas)」、同若手研究(B)「状況恐怖記憶における海馬-扁桃体経路の役割(研究代表者:牧野祐一)」による支援を受けて行われました。
原論文情報
Yuichi Makino, Denis Polygalov, Federico Bolanos, Andrea Benucci, and Thomas J. McHugh, “Physiological Signature of Memory Age in the Prefrontal-Hippocampal Circuit”, Cell Reports, 10.1016/j.celrep.2019.11.075
発表者
理化学研究所
脳神経科学研究センター 神経回路・行動生理学研究チーム
チームリーダー トーマス・マックヒュー(Thomas J. McHugh)
基礎科学特別研究員(研究当時) 牧野 祐一(まきの ゆういち)
報道担当
理化学研究所 広報室 報道担当
を用いた心肺蘇生が除細動不成功であったとしても…")
