
穂発芽しにくい農作物の効率的な開発に期待
2018-06-06 農研機構
ポイント
農研機構は、共同研究グループと共に、植物の種子休眠1)や乾燥ストレス応答2)において重要な働きをする植物ホルモン「アブシジン酸3)」が働くための新たな仕組みを明らかにしました。本成果により、穂発芽4)しにくい農作物の効率的な開発が期待されます。
概要
植物は乾燥や低温などのストレスにさらされると、植物ホルモンの一つであるアブシジン酸(ABA)を蓄積し、ABAが気孔の閉鎖や様々な遺伝子の発現量を調節することで、その環境に耐えることが知られています。また植物の種子は、発芽しても生きていけない環境では発芽しないで、種子休眠といわれる”種子を発芽させない”状態を維持しますが、ABAはこの種子休眠においても重要な役割を果たします。
今回農研機構は、共同研究グループと共に、発芽時にABAが働くための新たな仕組みを、モデル植物のシロイヌナズナで明らかにしました。これまでは発芽時にABAが働くには、ABA受容体5)を介した仕組みが必要と考えられていました。今回これ以外に、タンパク質脱リン酸化酵素タイプ2C(PP2C)6)のAHG17)タンパク質と、種子休眠で重要な働きをするDOG18)タンパク質の両方を介する新たな仕組みがあることを発見しました。
AHG1タンパク質やDOG1タンパク質の遺伝子は、イネやコムギ、オオムギなどの重要な作物にも存在しています。コムギやオオムギの栽培で問題となっている穂発芽は、種子休眠性に深く関わっていることから、ABA受容体を介した仕組みで働くタンパク質に加え、本研究で発見したAHG1タンパク質やDOG1タンパク質の機能を制御することで、穂発芽しにくい農作物の効率的な開発に役立つと期待されます。
本成果は、国際科学雑誌「Nature Communications」(2018年6月6日発行) のオンライン版に掲載されます。
関連情報
予算:科研費若手A (23688044, 26712030)、 科研費新学術研究(23119521)
お問い合わせ
研究推進責任者
農研機構次世代作物開発研究センター 所長 矢野 昌裕
研究担当者
農研機構次世代作物開発研究センター 基盤研究領域 西村 宜之
農研機構高度解析センター 生体高分子解析チーム 山崎 俊正、土屋 渉
岡山大学資源植物科学研究所 平山 隆志
広報担当者
農研機構次世代作物開発研究センター 広報プランナー 大槻 寛
詳細情報
研究の背景と経緯
植物は移動することができません。そのため、植物は乾燥や低温などのストレスにさらされると、植物ホルモンの一つであるアブシジン酸(ABA)を蓄積し、気孔の閉鎖や様々な遺伝子の発現量を調節することで、その環境に耐えることが知られています。また、植物の種子は、発芽しても生きていけない環境では発芽しないで、種子休眠といわれる”種子を発芽させない”状態を維持します。ABAはこの種子休眠においても、重要な役割を果たしており、種子休眠はコムギなどの重要な作物で問題になっている穂発芽性に深く関わっていることが知られています。
近年、世界人口の増加等による食料需要の増大や気候変動に起因する食料生産の不安定化が懸念されています。ABAが働くための仕組みや、そこで働く物質の役割を理解することができれば、得られた知見を応用して、気候変動による乾燥ストレスや異常な降雨などの劣悪環境下でも安定した収量を示す農作物の開発が可能になると期待されます。
ABAが働くための仕組みでは、タンパク質脱リン酸化酵素タイプ2C(PP2C)が重要な働きをするタンパク質として知られています。PP2Cは、ABI1とAHG1を各々含む、大きく2つのサブファミリーに分けることができます(図1a)。これまでに私たちを含む研究グループは、ABI1タンパク質は、ABA受容体であるPYR1タンパク質と結合し、このPYR1がABI1のPP2C活性を制御していることを明らかにしました(図1b)。一方、発芽時のABAが働くための仕組みでは、AHG1タンパク質が主要な役割を担っていることを以前報告していましたが、ABI1と同じPYR1を介した仕組みか、それとも別の仕組みで働いているかは分かっていませんでした。
研究の内容・意義
本研究では、AHG1タンパク質が働くための仕組みを明らかにすることを目的とし、以下の研究に取り組みました。
1)モデル植物であるシロイヌナズナを用いて、AHG1に結合するタンパク質を探索しました。その結果、ABA受容体であるPYR1タンパク質は同定されず、機能の詳細は分からないものの種子休眠で重要な働きをするDOG1タンパク質を同定することに成功しました。
2)DOG1タンパク質を過剰に蓄積する植物体(DOG1過剰発現体)は、通常であれば発芽可能な低い濃度のABA存在下において発芽や子葉が展開しにくいことから、ABA応答が高まっていることが分かりました(図2a)。次に、PP2Cタンパク質とDOG1との結合を調べたところ、DOG1はAHG1サブファミリーのPP2Cとは結合しますが、ABI1サブファミリーのPP2Cとは結合しないことを見出しました。さらに、AHG1とDOG1の関係を試験管内実験で調べたところ、DOG1がAHG1のPP2C活性を制御していることを明らかにしました(図2b)。
3)DOG1タンパク質の機能に関する成果としては、ヘム9)と呼ばれる二価の鉄原子とポルフェリンで構成される錯体と結合することによって赤褐色を示すことを見出しました(図3a)。ヘムとの結合には、DOG1タンパク質の245番目と249番目のアミノ酸残基であるヒスチジンが重要で、この2つのアミノ酸残基を別のアミノ酸に置き換えると、ヘムと結合できなくなりました(図3a)。また、この2ヶ所のアミノ酸に変異を導入したDOG1H245AH249A過剰発現体は、低濃度のABA存在下で発芽や子葉が展開しにくいというDOG1過剰発現体の性質が見られなくなることから、これまで機能が良く分かっていなかったDOG1が機能するためには、ヘムとの結合が重要であることを明らかにしました(図3b)。
以上より、これまでABA受容体であるPYR1タンパク質を介して制御されていると考えられていたPP2Cタンパク質のうち、AHG1サブファミリーのPP2CはDOG1タンパク質を介した仕組みで働くことを発見し、発芽を制御する仕組みの理解を飛躍的に進めることができました(図4)。
今後の予定・期待
近年、地球温暖化に起因する気候変動が世界各地の農作物の収量に負の影響を与えています。今後は、本研究で発見したAHG1タンパク質やヘムを介したDOG1タンパク質の機能を制御することで、穂発芽しにくい農作物の効率的な開発を進め、農作物の収量増加に貢献できると考えています。
用語の解説
1)種子休眠
種子が発芽に適切な時期が来るまで種子を発芽させない現象です。乾燥した種子は悪い環境にも強く、千葉市で発見された「大賀ハス」の種子は、2000年以上も種子休眠の状態を維持した後、発芽しました。
2)乾燥ストレス(応答)
植物が乾燥により水分を失った際に生じるストレスです。乾燥ストレス応答には、ABAを介する経路と介さない経路が存在することが知られています。
3)アブシジン酸(ABA)
植物ホルモンの一つで、植物の乾燥などのストレス応答で働く重要な調節物質であり、種子休眠や種子発芽の抑制、気孔の閉鎖など多岐にわたり作用することが知られています。近年、ABA受容体が同定され、主要なABAが働くための仕組みが明らかになりました。
4)穂発芽
作物の収穫前に、穂に着生している種子が、降雨などにより発芽してしまう現象です。オオムギやコムギなどでは、発芽による品質の低下で商品価値が失われることから、農業上の大きな問題となっています。
5)ABA受容体
ABAと結合し、ABAの作用を起こすために働くタンパク質です。これまでにABA受容体の一つとしてPYR1タンパク質が同定されており、ABA受容体であるPYR1は複数のPP2Cと結合して、それらのPP2C活性を直接制御することが知られています。
6)タンパク質脱リン酸化酵素タイプ2C (PP2C)
タンパク質脱リン酸化酵素の一つで、リン酸化されたタンパク質のリン酸基を脱リン酸化する酵素タンパク質です。真核生物に広く保存され、植物はヒトなどと比べると非常に多くのPP2Cが存在します。PP2Cタンパク質はABAが働くための仕組みで重要な役割を担い、シロイヌナズナではABI1やAHG1などが知られています。
7)AHG1
低濃度のABA存在下で発芽することができない突然変異体の原因遺伝子として同定され、その原因遺伝子はPP2Cの一つでした。主に種子で働いていると考えられています。
8)DOG1
種子休眠の重要な量的形質遺伝子座として同定されました。DOG1タンパク質の配列は、これまでに機能が知られているタンパク質の配列と相同性が無いため、DOG1の機能はよく分かっていませんでした。
9)ヘム
二価の鉄原子とポルフェリンで構成される錯体です。タンパク質と結合するものが多く、その代表的なものとして、ヘムとグロブリンから成るヘモグロビンが知られています。ヘモグロビンは、ヘムの鉄原子が酸素分子と結合することで、酸素を運搬しています。
発表論文
Noriyuki Nishimura, Wataru Tsuchiya, James J. Moresco, Yuki Hayashi, Kouji Satoh, Nahomi Kaiwa, Tomoko Irisa, Toshinori Kinoshita, Julian I. Schroeder, John R. Yates, III, Takashi Hirayama & Toshimasa Yamazaki, “Control of seed dormancy and germination by DOG1-AHG1 PP2C phosphatase complex via binding to heme”, Nature Communications 2018 (doi: 10.1038/s41467-018-04437-9)
参考図表

図1. シロイヌナズナのABA応答に働くPP2Cの分子系統樹と既知のABA応答のモデル図
a) ABA応答に働くPP2Cは、大きくABI1サブファミリーとAHG1サブファミリーの2つに分けることができます。
b) ABAと結合したABA受容体であるPYR1は、ABI1と結合し、ABI1のPP2C活性を抑制(減少)することで、植物はABA応答します。
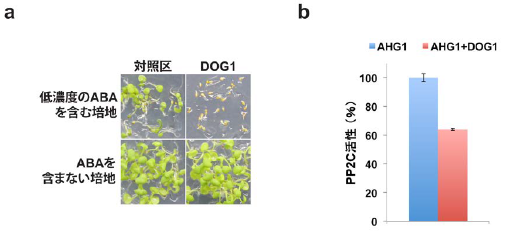
図2. ABA応答に働くDOG1タンパク質の役割
a) ABA応答に働くPP2Cは、大きくABI1サブファミリーとAHG1サブファミリーの2つに分けることができます。
b) ABAと結合したABA受容体であるPYR1は、ABI1と結合し、ABI1のPP2C活性を抑制(減少)することで、植物はABA応答します。
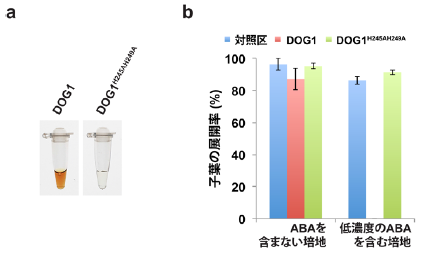
図3. DOG1タンパク質はヘムと結合する
a) DOG1タンパク質は、ヘムと結合するため赤褐色を示します(左)。しかし、ヘムと結合する245番目と249番目のアミノ酸残基であるヒスチジン(H)をアラニン(A)に置き換えた変異導入DOG1タンパク質(DOG1H245AH249A)は、ヘムと結合できなくなるため、赤褐色を示しませんでした(右)。
b) DOG1過剰発現体(赤バー)は、対照区(青バー)と比べ、低濃度のABA存在下で発芽や子葉が展開しにくいです(子葉の展開率はほぼ0%)。しかし、ヘムと結合する2ヶ所のアミノ酸に変異を導入したDOG1H245AH249A過剰発現体(緑バー)では、対照区と同様に、低度濃度のABA存在下においても発芽や子葉が展開します。
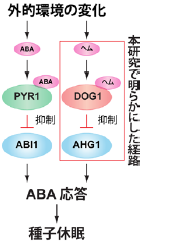
図4. 本研究から考えられる発芽の制御に関わるモデル図
発芽を制御するABA応答は、既に知られているABA受容体PYR1を介する経路(左側)と本研究で明らかにしたDOG1を介する経路(右側)があります。外的環境の変化により、PYR1やDOG1はABAやヘムなどの調節物質と結合し、それらがABI1やAHG1などのPP2C活性をそれぞれ制御することで、ABA応答が誘導され、種子は休眠すると考えられます。
