2022-12-24 東京大学
発表のポイント
- セルロースを酸化的に分解する酵素「溶解性多糖モノオキシゲナーゼ(LPMO)」が、どのように他の酵素の反応性を高めるのかを調べました。
- 結晶性セルロース表面のLPMOによって酸化されたところの周辺で、非晶化の連鎖反応が起こり、他の酵素による分解性が高まることが明らかになりました。
- 持続可能でカーボンニュートラルな社会の構築を達成するためには、地球上で最も豊富に存在するバイオマスであるセルロースを、様々な用途に利用していく必要があります。その鍵となる酵素によるセルロース分解機構の詳細が明らかになったことで、効率良くバイオ化成品やバイオ燃料、バイオプラスチックなどを生産することが可能となります。
発表概要
きのこが生産するセルラーゼ(Cel6AとCel7D)による結晶性セルロースの分解が、溶解性多糖モノオキシゲナーゼ(LPMO)の一種であるAA9Dの添加によって促進されることが、生化学的実験で明らかにされました。その機構を、高速原子間力顕微鏡(注1)による酵素分子の観察、および分子動力学シミュレーション(注2)によって調べたところ、AA9Dが酸化的に切れ込み(ニック)を入れると、反応したセルロース分子だけでなく、その周辺のセルロース分子も連鎖的に水和、非晶化し、他の酵素による分解性を高めていることが分かりました。この知見から、地球上で最も豊富に存在するバイオマスであるセルロースからバイオ化成品やバイオ燃料、バイオプラスチックなどを生産する効率を著しく高めることが可能となり、循環型で生物圏に優しい「サーキュラーバイオエコノミー」の達成に貢献することが期待されます。
発表内容

図1 きのこの一種P. chrysosporium由来のLPMO(AA9D、橙)によって酸化された結晶性セルロースの表面を分解するセロビオヒドロラーゼ(Ce6A、黄 Cel7D、桃)。
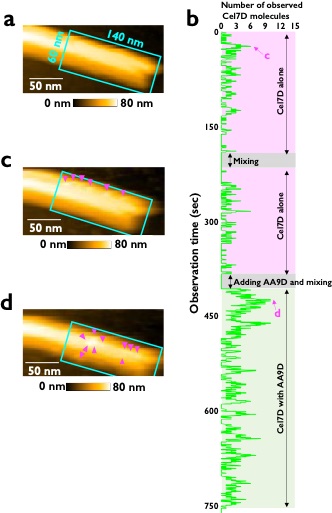
図2 高速原子間力顕微鏡によるセルラーゼ分子の観察。
a:酵素を入れる前の結晶性セルロース、b:酵素分子の時間変化、c:Cel7Dのみが反応している時のセルロース表面の様子、d:Cel7DとAA9Dが一緒に反応している様子。明るい粒子がセルラーゼ分子で、AA9Dが存在しているときに観察されるセルラーゼ分子の数が増えている様子が分かる。
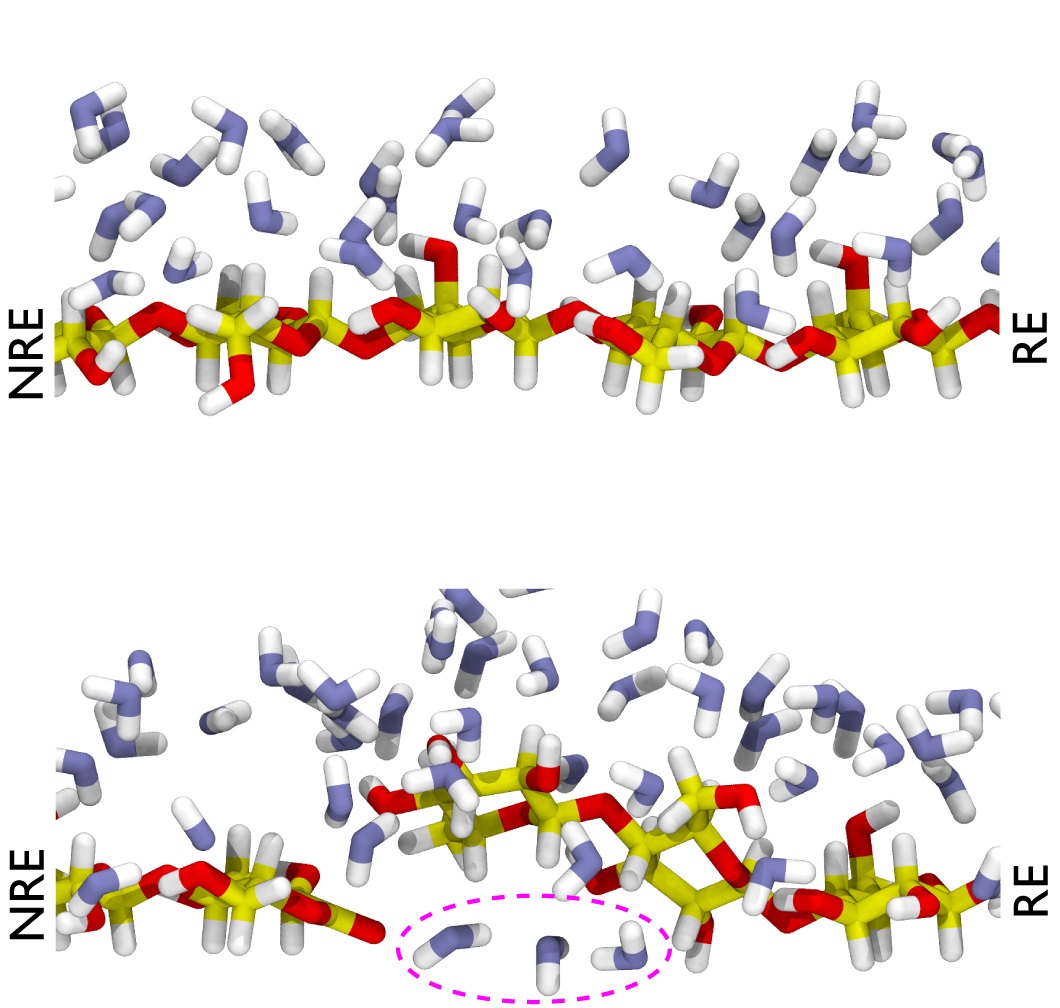
図3 AA9Dによって切断された末端がラクトンになっている部位周辺における水分子の分子動力学シミュレーション結果。
上段:結晶性セルロースの表面(コントロール)。
下段:AA9Dによって切断された後の結晶表面。点線で囲われているのはセルロース分子の下に流れ込んだ水分子。
RE:還元性末端、NRE:非還元性末端
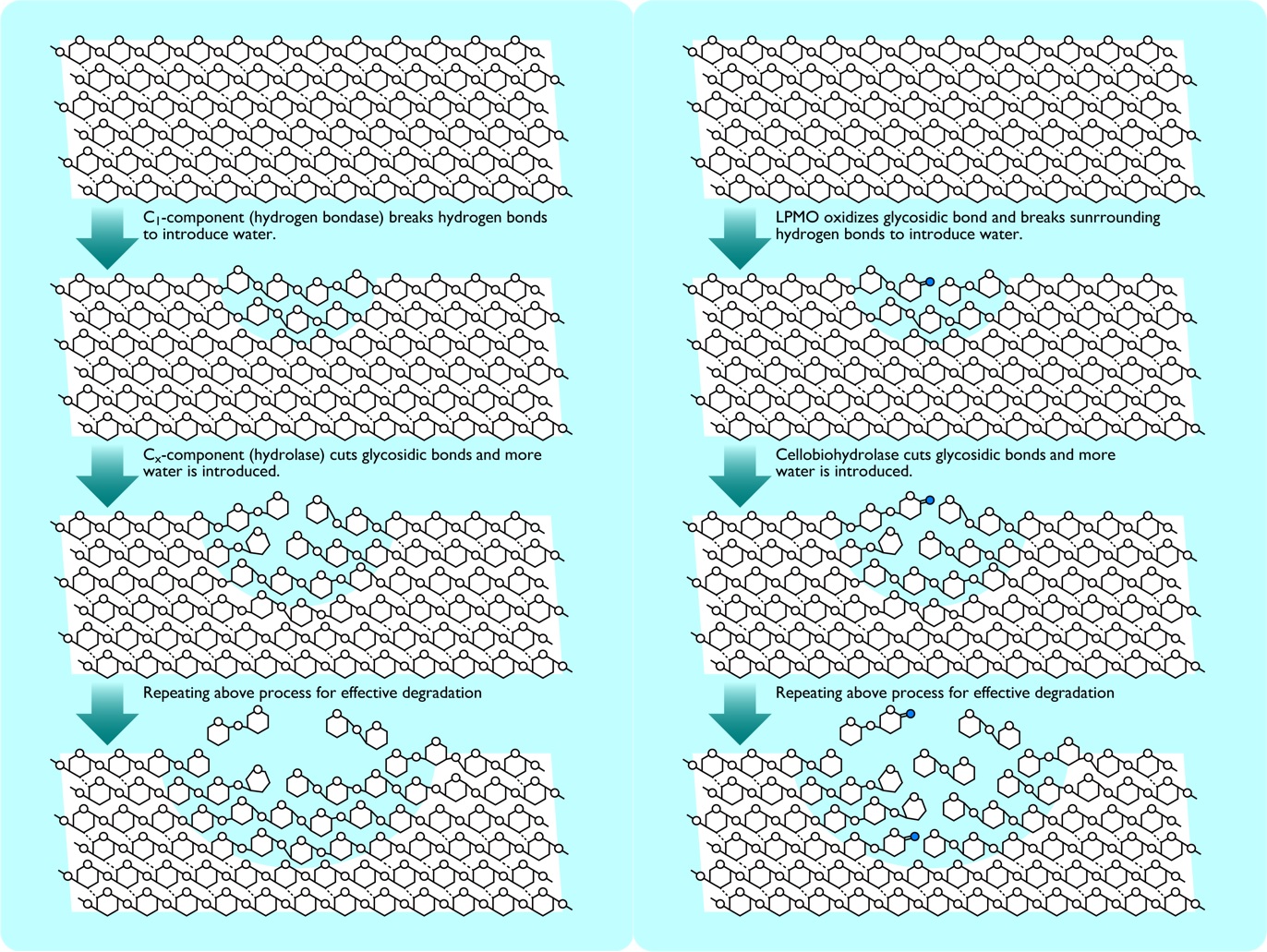
図4 C1-Cx説(左)およびLPMOによるoxidative boost(右)におけるセルロース分子の分解。
LPMOによって酸化された箇所(青丸)周辺に水が流れ込んで非晶化する過程は、C1-Cx説とほとんど変わらないことがわかる。
セルロースは植物細胞壁の主成分であり、地球上で最も豊富に存在するバイオマスです。セルロースはグルコースが数千から数万繋がった分子であることから、効率良く分解することができれば、得られたグルコースからバイオ燃料やバイオマスプラスチック等様々な化合物を作ることが可能になります。しかしながら、セルロースは水に溶けにくい物質であることから、同じくグルコースがいくつも繋がってできている澱粉(でんぷん)と比較しても、水を使った分解(加水分解)が非常に難しいという欠点があります。その一方で、自然界にはきのこやカビ、バクテリアのように難分解性のセルロースを効率良く分解し、成長するためのエネルギー源としている生き物がおり、それらの生き物は全てセルロース分解酵素(セルラーゼ)を使ってセルロースを消化して栄養源としていることが知られています。
酵素によるセルロースの分解に関する研究は、古くは19世紀後半から行われていましたが、その機構に関してはじめて論じられたのは1950年のReeseらによる報告でした。その論文でReeseらは、セルロースははじめにC1と呼ばれる成分によってグルコース同士の結合が切られることなく水和し、水和したセルロースがCx成分と呼ばれる酵素によって加水分解されるというメカニズム「C1-Cx説」を提唱しました(参考文献1)。しかしながら、C1成分に相当する酵素はその後20年経っても発見されず、その間に植物細胞壁中のセルロースには結晶と非晶(結晶が乱れて水和しやすくなった構造)があることが知られるようになり、1972年にはendo型の酵素が非晶性のセルロースを分解し、そこからexo型のセルラーゼが結晶性セルロースを壊していく「endo-exo説」が提唱され(参考文献2)、その後広く受けいれられるようになりました。一方で、五十嵐圭日子教授が学生時代に師事していたこともある米国ジョージア大学の故Eriksson教授らは、窒素もしくは酸素を溶かした溶液中できのこやカビから抽出されたセルロース分解酵素を反応させたところ、酸素を溶かした溶液でセルロース分解が速く進むことを1974年に発見し、酸化反応がセルロース分解を速める「oxidative boost(酸化的加速)」を提唱しました(参考文献3)。その後、セロビオース脱水素酵素(CDH)と呼ばれる酵素がその原因酵素であるとして研究が進められていましたが、そのメカニズムに関しても現在に至るまで不明なままでした。
そのような状態が長く続いている中で、2010年にノルウェーの研究チームが「oxidative boost」の原因酵素として「溶解性多糖モノオキシゲナーゼ(LPMO)」という酵素を発見しました(参考文献4)。LPMOは分子内に銅原子を持っており、結晶性のセルロースを酸化的に開裂させ、そこからその他の酵素が反応するという仕組みで加速していると考えられました。その後、CDHがLPMOの銅原子を還元できることが分かり、これらの酸化還元酵素がoxidative boostに関与する機構が提唱されました。
五十嵐教授の研究チームは、セルロースの中でも特に分解性が低い結晶性セルロースの酵素による分解を長年研究しており、きのこの一種であるPhanerochaete chrysosporiumが生産するLPMOのAA9Dがその他のセルロース分解酵素(Cel6A、Cel7D)による結晶性セルロースの分解にどのような影響を与えるのかに関して実験を行っていました。その過程で、AA9Dが自身の生成物である酸化されたオリゴ糖をほとんど出さずに、その他の酵素の活性を高めることを発見しました。そこで、名古屋大学の内橋貴之教授の研究チームとの共同研究で、セルロース表面で働くCel7D分子の様子を高速原子間力顕微鏡によって観察したところ、AA9Dが一緒に働いている時は、セルロース表面を動く酵素分子の数が多くなることが明らかになりました。さらに、米国エネルギー省再生可能エネルギー研究室のGregg T. Beckham博士の研究チームとの共同研究によって、スーパーコンピュータを用いてAA9Dによって酸化されたセルロース表面がどのように変化するのかを分子動力学シミュレーションしたところ、AA9Dによって酸化されたセルロースの表面では、酸化されたセルロース分子が浮いてくるだけでなく、周りのセルロース分子との水素結合も切れて、そこに水分子が流れ込んで溶ける(非晶化する)ことが分かりました。
以上の結果を全て統合して考えると、LPMO(本研究ではAA9D)が一度セルロースを酸化的に開裂させると、その周辺の水素結合ネットワークが乱されて、酸化されていないセルロースも水和して非晶化し、その結果他のセルロース分解酵素(本研究ではCe6AとCel7D)が非晶化されたセルロース分子を容易に分解できるようになり、それらの酵素が反応するとさらにその周辺のセルロースが水和して非晶化されるという連鎖的な反応が始まることで「oxidative boost」が起こっていると考えられました。このように非晶化したセルロースを分子の途中から切り始める機構は「endo-exo説」で提唱された仕組みと同じであるのと同時に、このように「水和」から始まるドミノ倒しのようなセルロース分解は、Reeseらが1950年に提唱した「C1-Cx説」と同じ仕組みです。すなわち、これら3つの学説、C1-Cx説、endo-exo説、oxidative boostは、同じ現象を違う視点で見ていただけであることになり、70年にわたる議論に終止符を打つ決定打となりました。
本研究成果は、木や草などの主成分であるセルロースを効率良く酵素で分解するために直接応用することが可能であることから、稲わらやサトウキビバガス(サトウキビの絞りかす)等の未利用バイオマスや、古紙や建築廃材などのセルロース系のバイオマスから、バイオ化成品やバイオ燃料、バイオプラスチック等を生産するために重要な知見となります。
なお、本研究成果は、米国科学振興協会(AAAS)が発刊しているオープンアクセス雑誌「Science Advances誌」に掲載されます。
本研究は、文部科学省科学研究費補助金新学術領域研究(18H05484)「植物の力学的最適化戦略に基づくサステナブル構造システムの基盤創成」(計画研究代表者:五十嵐圭日子、領域代表:奈良先端科学技術大学 出村 拓 教授)、基盤研究(B)(19H03013)「きのこのゲノム編集技術を利用した木材腐朽現象の理解とバイオマス変換系の構築」(研究代表者:五十嵐圭日子)、(21H01772)「機械刺激を加えた単一タンパク質の構造および力学特性応答の高速AFM解析」(研究代表者:内橋貴之)、基盤研究(C)(15K07383)「放線菌由来セルラーゼの結晶性基質の効率的分解に関わる機構解明」(研究代表者:内山拓)、環境省・(独)環境再生保全機構(ERCA)環境研究総合推進費(JPMEERF21S11902)「プラスチックの持続可能な資源循環と海洋流出制御に向けたシステム構築に関する総合的研究」(サブテーマ研究代表者:五十嵐圭日子、プロジェクトリーダー:東北大学 吉岡敏明 教授)、およびBusiness Finlandフィンランド卓越教授(FiDiPro)プログラム「Advanced approaches for enzymatic biomass utilization and modification (BioAd)」(招聘教授:五十嵐圭日子)の補助を受けたものです。
発表者
- 内山 拓(東京大学大学院農学生命科学研究科生物材料科学専攻 (研究当時))
- 内橋貴之(東海国立大学機構 名古屋大学理学研究科、糖鎖生命コア研究拠点(iGCORE)教授/自然科学研究機構生命創成探求センター 客員教授)
- 石田卓也(東京大学大学院農学生命科学研究科生物材料科学専攻 博士研究員(研究当時))
- 中村彰彦(分子科学研究所生命・錯体分子科学研究領域 クロスアポイントメント准教授/静岡大学農学部応用生命科学科 テニュアトラック准教授)
- Josh V. Vermaas(ミシガン州立大学生化学分子生物学科 助教)
- Michael F. Crawly(米国エネルギー省再生可能エネルギー研究室 センターディレクター)
- 鮫島正浩(東京大学大学院農学生命科学研究科生物材料科学専攻 東京大学名誉教授/信州大学特任教授)
- Gregg T. Beckham(米国エネルギー省再生可能エネルギー研究室 シニアリサーチフェロー)
- 五十嵐圭日子(東京大学大学院農学生命科学研究科生物材料科学専攻 教授/VTTフィンランド技術研究センター 客員教授(研究当時))
発表雑誌
- 雑誌名
- Science Advances
- 論文タイトル
- Lytic polysaccharide monooxygenase increases cellobiohydrolases activity by promoting decrystallization of cellulose surface
- 著者
- Taku Uchiyama, Takayuki Uchihashi, Takuya Ishida, Akihiko Nakamura, Josh V. Vermaas, Michael F. Crowley, Masahiro Samejima, Gregg T. Beckham, and *Kiyohiko Igarashi(*責任著者)
- DOI番号
- 10.1126/sciadv.ade5155
- 論文URL
- https://www.science.org/doi/10.1126/sciadv.ade5155
問い合わせ先
東京大学大学院農学生命科学研究科 生物材料科学専攻
教授 五十嵐 圭日子(いがらし きよひこ)
参考文献
- Reese E. T., Sui R. G. H. & Levinson H. S., The biological degradation of soluble cellulose derivatives and its relationship to the mechanism of cellulose hydrolysis, J. Bacteriol. 59:485-497 (1950)
- Wood T. M. & McCrae S. I. The purification and properties of the C1 component of Trichoderma koningii cellulase. Biochem. J. 128:1183–1192 (1972)
- Eriksson K.-E., Pettersson B., & Westermark U., Oxidation: An important enzyme reaction in fungal degradation of cellulose, FEBS Lett. 49:282-285. (1974)
- Vaaje-Kolstad G., Westereng B., Horn S. J., Liu Z., Zhai H., Sørlie M., Eijsink V. G., An oxidative enzyme boosting the enzymatic conversion of recalcitrant polysaccharides. Science 330:219-222 (2010)
用語解説
注1 高速原子間力顕微鏡
原子間力顕微鏡は、探針(プローブ)を観察対象の表面に沿って走査することで観察対象の形の情報(画像)を得ます。しかしながら従来の原子間力顕微鏡は1画像取得するのに数分を要していたため、対象物の変化をリアルタイムで追うことは困難でした。高速原子間力顕微鏡では、様々な改良を行うことで走査の高速化に成功し、リアルタイムで画像を撮ることができます。本研究ではセルラーゼ分子がセルロース表面でどのように振る舞っているのかを観察するために用いられています。
注2 分子動力学シミュレーション
物質を構成する各原子に対して、古典力学におけるNewtonの運動方程式を解き、計算機(コンピュータ)によって原子位置やエネルギーの時間変化を追跡する手法。本研究では、高速原子間力顕微鏡では観察できない水分子の挙動を推測するために用いられています。

