
2022-08-04 東京大学
1. 発表者:
大木 研一(東京大学 大学院医学系研究科機能生物学専攻 教授/Beyond AI研究推進機構 教授/国際高等研究所ニューロインテリジェンス国際研究機構 主任研究者(WPI-IRCN))
村上 知成(東京大学 大学院医学系研究科機能生物学専攻 助教)
2. 発表のポイント:
◆複雑な脳神経ネットワークを効率的に形成するための新たなメカニズムを明らかにした。
◆マウス大脳視覚野と視床核を含む領野間結合がどのように形成されるかを網羅的に調べた世界初の研究である。
◆将来的に先天性盲などの疾患に対する治療法や、優れた人工知能を形成するための回路形成アルゴリズムに応用されることが期待される。
3.発表概要:
東京大学大学院医学系研究科機能生物学専攻統合生理学分野の村上知成助教、大木研一教授、松井鉄平講師(研究当時、現:岡山大学大学院自然科学科 准教授)らの研究グループは、大脳視覚野にある多数の領野間結合が、発達時に効率的に形成されるメカニズムを解明した。
ヒトの大脳皮質には180にも及ぶ多数の領野が存在し、これらの領野間を結ぶ数千にも及ぶ精密な神経回路による階層的かつ並列的な情報処理は、私たちの脳が複雑かつ汎用的な知性を獲得するための基盤となっている。従来の神経回路発達の研究では、感覚器から大脳皮質の入り口まで(視覚の場合、網膜から一次視覚野まで)の神経回路形成については詳細に調べられてきたが、大脳皮質の領野間をつなぐ無数の結合がどのようなメカニズムで3次元の脳内で精密に混線なく配線されるのかについてはほとんど分かっていなかった。そのメカニズムの一つとして領野の階層性に従ってより低次な領野からより高次な領野へと順に形成していけば混線なく配線できると考えられるが、マウスでは生後から開眼までの2週間の間に多数の領野間の結合を全て作る必要があり、このメカニズムでは時間がかかり過ぎる。それでは、このような複雑な無数の領野間結合を短時間に混線なく形成するためのメカニズムは何だろうか。
本研究ではマウスの視覚系を用いて、大脳皮質の多数の視覚関連領野間をつなぐ結合が短期間で効率的に形成されるメカニズムを解明した。まず、領野間結合が形成される前に、網膜と大脳皮質の多数の視覚関連領野をつなぐ経路が先に形成されることを見いだした。さらに、この経路を伝播する網膜由来の自発活動により、網膜の場所をあらわす情報が多数の視覚関連領野に伝えられ、これが教師信号となって、大脳皮質の多数の視覚関連領野の網膜座標が対応する場所を精密に結ぶ結合が形成されることが示された。
本研究は大脳皮質の領野間結合の形成メカニズムを明らかにしただけでなく、先天盲や早期失明の病態を理解する上でも重要であり、また汎用的人工知能の開発に寄与する可能性が考えられる。本研究成果は、2022年8月3日(英国夏時間)に英国科学誌「Nature」のオンライン版に掲載されました。
本研究は、Beyond AI研究推進機構、日本医療研究開発機構(AMED)による「脳とこころの研究推進プログラム(革新的技術による脳機能ネットワークの全容解明プロジェクト)、ニューロインテリジェンス国際研究機構(IRCN)、科研費「基盤研究S(課題番号:25221001、19H05642)」「学術変革領域研究(A)(課題番号:20H05917)」「学術変革領域研究(B)(課題番号: 21H05165)」「若手研究(課題番号:21K15181)」、JST さきがけ(JPMJPR19M9)の支援を受けて実施されました。
4.発表内容:
<研究の背景>
階層的かつ並列的な視覚ネットワーク(注1)は、霊長目、げっ歯目、食肉目と種を超えて保存されており、視覚情報処理の基本構造と考えられている(図1)。発達期に我々の脳は、この複雑な視覚ネットワークを遺伝子発現と自発活動を含む神経活動の両方の助けを借りて形成している。神経ネットワークの発生過程は重要なテーマとして以前から盛んに研究が行われていたが、どの感覚系においても末梢から視床核(注2)を介して大脳一次感覚野に至るまでの低次経路に留まっており、大脳皮質の高次領野や他の視床核を含む神経ネットワーク全体がどのように形成されるかほとんど調べられていなかった。
神経ネットワーク形成を解明するためには、大脳皮質視覚野と視床核を含む視覚ネットワーク全体において、領野間の結合がどのタイミングで形成されるかを追跡する必要がある。しかしながら、発達期の未熟な脳においてそれぞれの領野の正確な位置を特定することは困難であり、技術的なボトルネックとなっていた。本研究では、発達期の大脳皮質で観察される自発活動パターンを詳細に解析することで、未熟なマウス大脳皮質での一次視覚野(V1)と多数の高次視覚野の位置を正確に同定できることを発見した。この方法を用いて、マウス視覚ネットワーク全体で領野間結合がどのように形成されるのか、さらに観察される自発活動が領野間結合形成にどのような機能的役割を果たすかを調べた。
<研究内容>
まず我々は開眼前の未成熟な状態の遺伝子改変マウス(注3)を用いて、大脳皮質視覚野全体の自発的神経活動を広視野カルシウムイメージング(注4)により観察した。自発活動の時空間的なパターンを機能的相関解析(注5)したところ、生後5日というまだ領野間結合が形成される前から、各高次視覚野がレチノトピー構造(注6)を持つことを発見した。このレチノトピー構造からV1と複数の高次視覚野の正確な位置を同定することができ、領野間結合の発達を追跡することが可能となった。
我々は同定した領野間結合の発達を解剖学的に調べるために、自発活動の解析から同定した一つの視覚野にニューロトレーサー(注7)であるカルボシアニン色素(DiI、注8)を注入し、他の領野で逆行性(注9)に染まる細胞を観察した。その結果、視床核から大脳皮質視覚野への投射[低次視床核→V1、高次視床核→高次視覚野]が、大脳皮質の領野間結合よりも先に形成されることを発見した。さらに低次・高次視床核はそれぞれ独立の経路で網膜から入力を受けており、網膜からV1と高次視覚野まで並列な経路(モジュール)として形成されていることを明らかにした(図2)。また、我々は解剖学的手法だけでなく、薬理学的な抑制実験(注10)を用いて、これらの並列モジュールは網膜からの自発活動を大脳皮質まで伝播していることも明らかにした。これによりV1だけでなく高次視覚野がレチノトピー構造を早期から持ち、大脳皮質の領野間結合はこのレチノトピー構造を元にして効率的に形成していることが示唆された。最後に我々は並列モジュールを伝播する網膜自発活動の、大脳皮質の領野間結合の形成に対する役割を調べるために、産まれた直後から網膜自発活動を除去し、V1から高次視覚野への投射形成における影響を調べた。その結果、網膜自発活動を除去したマウスではV1から高次視覚野への投射が乱れており、このことは網膜自発活動が大脳皮質の領野間結合の形成に重要であることを示している。今回の研究は脳神経ネットワークが形成される発達過程を示しただけでなく、神経活動、遺伝子発現パターンに因る形成メカニズムを解明するための基盤となることが期待される。
<今後の展望>
本研究成果は将来的に先天性盲などの疾患治療の分野への応用が期待される。全世界で視力障害者は2億人上、失明者は3600万人にのぼる。先天盲や早期失明の患者では、点字解読により高次視覚野やV1が活動することが報告されているが、その解剖学的基盤については、ほとんど知られていない。本研究では早期失明によりV1から全ての高次視覚野への結合が著しく減少し、網膜座標系を特異的につなぐ結合も見られなくなることを見出しており、高次視覚野は他の領野から入力を得ている可能性が示唆される。このような早期失明による大脳皮質の領野間結合の可塑的変化を調べることは、先天盲や早期失明の病態を理解する上で重要と考えられる。
また、本研究成果は人工知能研究の分野に対しても重要な知見を示唆している。現在の人工知能は多数の階層をもった深層学習の方向へと発展してきている。並列分散処理は複数のチャンネルを使うことにより一部取り入れられているものの、形と動き、顔と建物のように、大きく異なる特徴量を明示的に並列処理するようには作られていない。大脳皮質では、階層的処理に加えて並列分散処理は大きな原理の一つであり、それにより私たちの脳は複雑かつ汎用的な知性を獲得できていると考えられる。上記の結果は、この並列分散処理を担う回路が、網膜と大脳皮質をつなぐ従来知られていなかった回路を伝播する情報を教師信号として、形成されることを示したものであり、この原理を人工知能に応用し、階層処理だけでなく並列分散処理も取り入れることにより、汎用的な人工知能の開発に役立つ可能性が考えられる。
5.発表雑誌:
雑誌名:「Nature」(オンライン版:8月3日)
論文タイトル:Modular strategy for development of the hierarchical visual network in mice
著者:Tomonari Murakami*, Teppei Matsui*, Masato Uemura, Kenichi Ohki*
DOI番号:10.1038/s41586-022-05045-w
アブストラクトURL:www.nature.com/articles/s41586-022-05045-w
6.問い合わせ先:
<研究に関すること>
東京大学大学院医学系研究科 機能生物学専攻 統合生理学分野
教授 大木 研一(おおき けんいち)
<広報に関すること>
東京大学医学部・医学系研究科 総務チーム
東京大学国際高等研究所 ニューロインテリジェンス国際研究機構 広報担当(WPI-IRCN)
東京大学産学協創部(Beyond AI研究推進機構 広報担当)
7.用語解説:
注1 階層的かつ並列的な視覚ネットワーク
大脳皮質視覚野は霊長類で30、齧歯類でも10領野以上存在し、これらが階層的かつ並列的に結合することで視覚情報処理が行われている。大脳皮質の低次視覚野を出た後、頭頂葉へ向かう背側経路と側頭葉へ向かう腹側経路の2つに分かれ、それぞれ物の動き・形といった異なる視覚情報を並列に処理する。
注2 視床核
脳構造において大脳皮質の内側に位置し、間脳の一部を占める部位。視覚、聴覚、体性感覚などの感覚入力を大脳皮質の一次感覚野へ中継する低次視床核と、大脳皮質の高次領野と相互的な結合を持つ高次視床核がある。
注3 遺伝子改変マウス
人工的な操作により外来性の遺伝子を導入されたマウス、あるいは内在性の遺伝子に何らかの変異が導入されたマウス。本実験で用いた遺伝子改変マウスは、カルシウム感受性タンパクであるGCaMP6fを大脳皮質の興奮性細胞に発現するように、外来性の遺伝子を導入されたマウスである。
注4 広視野カルシウムイメージング
大脳皮質の神経活動を多数の領野を含む広域で観察する実験手法。神経細胞が活動し、細胞内のカルシウム濃度が上昇すると、カルシウム感受性タンパクGCaMP6fの緑の蛍光が明るくなる。本研究ではこの蛍光の明るさの時間的な変化を計測し、神経細胞の集団活動を観察した。
注5 機能的相関解析
各脳領域で観察される自発活動の相関係数を算出することで、自発活動の同期性を検証する解析法。自発活動がよく同期していれば相関係数が高くなり、そうでなければ低くなる。ある領野間の自発活動が同期しているということは、片方の領野で起こった活動がもう一つの領野へ直接的または間接的に伝播している、あるいは両方の領野へ別の領野の活動が伝播していることを示唆している。
注6 レチノトピー構造
各視覚関連領野において、視野の空間的情報が保持され表現される構造。個々の神経細胞は視野上の特定の位置からのみ入力を受けており、この視野上の位置は受容野と呼ばれる。各視覚関連領野には、この受容野が視野全体を網羅するように神経細胞が配置されている。これは、網膜からの神経回路が視床核を介して大脳の視覚関連領野へとつながるときに、上下左右の位置関係を保存したまま結合することによる。
注7 ニューロトレーサー
神経細胞の細胞体や軸索を色付けし、細胞の形態や領野間の結合を解剖学的に標識するために用いる薬剤あるいは蛍光タンパク質。
注8 カルボシアニン色素(DiI)
細胞膜を標識する脂溶性の色素。細胞体だけでなく軸索からも色素が伝播するため、ある領野へ投射を送る細胞について、投射先である軸索側から取り込ませれば、投射元である細胞体を染めることができる。
注9 逆行性
神経細胞に外来性の物質を導入したときに、細胞内で物質が伝播する方向を指す。細胞体から軸索へ伝播する場合が順行性、軸索から細胞体へ伝播する場合が逆行性である。
注10 薬理学的な抑制実験
薬剤投与により神経細胞の活動を局所的に抑制する実験手法。抑制性神経伝達GABAのアゴニストであるムシモルや、興奮性神経伝達に働く細胞膜上の電位依存性ナトリウムチャネルを抑制するテトロドトキシン等を脳内へ投与すると、薬剤が拡散する範囲において神経細胞の活動が抑制される。本実験で用いたものはムシモルである。
8.添付資料:

図1:マウス視覚神経経路の構造
左、マウス大脳皮質視覚野の模式図。
右、網膜から高次視覚野への階層的ネットワークの概要。高次視覚野は腹側(青)・背側経路(赤)に分かれており、視覚情報を並列に処理する。この階層的・並列な神経ネットワークは種を超えた哺乳類脳の基本構造であるが、発達期においてこの経路がどのように形成されるか未だ明らかとなっていない。
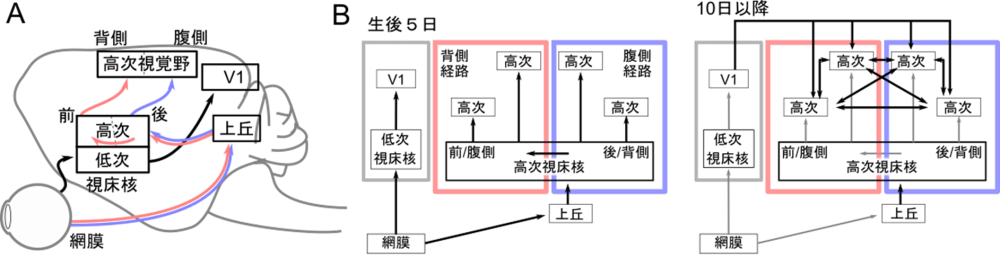
図2:本研究で明らかとなった並列モジュール式の発達戦略
A、生後5日で観察された視覚に関与する領野間の結合
異なる視床核を経由して、網膜からV1と高次視覚野への経路が並列に形成されている(並列モジュール経路)。
B、視覚経路の発達過程
生後5日では大脳皮質領野間の結合よりも前に、網膜からV1と高次視覚野への並列モジュール経路が形成される。その後10開港に並列モジュールを統合するために大脳皮質領野間の結合が形成され、階層的ネットワークが完成される。
")
