
2023-08-09 京都大学iPS細胞研究所
ポイント
- TJP1の発現抑制は、多能性幹細胞のBMP4に対する反応性を亢進させ、原腸陥入に類似した分化パターン形成を消失させる
- TJP1の発現抑制による多能性幹細胞のBMP4反応性の亢進は、細胞間接着のバリアー機能低下に由来する
- TJP1の発現抑制は、多能性幹細胞を始原生殖細胞に類似した細胞へと分化しやすくする
- TJP1の発現抑制は、多能性幹細胞のBMP4への反応性を増強することにより、始原生殖細胞様細胞への分化を促進する
1. 要旨
Ivana Vasic 氏(当時:カリフォルニア大学 博士課程大学院生、現:Vitra Labs Founder/CEO)、友田紀一郎准教授(CiRA未来生命科学開拓部門、グラッドストーン研究所 研究員)、山中伸弥教授(CiRA同部門、グラッドストーン研究所 上席研究員)ら、グラッドストーン研究所iPS細胞研究拠点(アメリカ)の研究グループは、細胞間接着注1)がヒト多能性幹細胞からの分化において重要な役割を果たし、細胞間接着の抑制が始原生殖細胞注2)への分化を促すことを発見しました。
グラッドストーン研究所iPS細胞研究拠点は、京都大学オンサイトラボ制度の一環として2019年に認定・設置されました。CiRA未来生命科学開拓部門の山中伸弥教授は、2007年からグラッドストーン研究所で上席研究員として研究室を主宰しています。
正常な体の組織は、初期発生にみられる原腸陥入注3)などのパターン形成イベント注4)によりかたち作られます。それらのイベントを制御する分子メカニズムを深く理解することは、個体発生の根幹や病気の原因を理解すること、また、再生医療のための細胞や組織を生体外で構築するために必要です。
本研究では、ヒト多能性幹細胞を分化誘導することにより再現した原腸陥入モデルを用いて、BMP4シグナル伝達による原腸陥入時のパターン形成における細胞間接着の役割を明らかにしました。
細胞間接着の一つであるタイトジャンクション(密着結合)注5)を制御する重要な分子TJP1注6)の発現を抑制したところ、BMP4シグナルに対する反応性が増し、原腸陥入に類似したパターン形成の消失がみられました。これらの結果から、TJP1がこの原腸陥入モデルにおいて重要な役割を担っていると考えられます。
さらに、研究チームはTJP1の発現抑制によって多能性幹細胞が始原生殖細胞(primordial germ cell: PGC)の特徴を示す細胞に著しく分化しやすくなることも見出しました。また、こうしたTJP1の発現抑制による効果の大部分は、細胞間接着分子の消失によってBMP4シグナル伝達への反応性が亢進したことに起因することを示しました。
これらの結果を発展させ、通常のヒト多能性幹細胞から、TJP1を抑制することなく、PGC様細胞へと効率よく分化させる方法を示しました。本研究は、培養細胞において分化誘導に関わるシグナル伝達機構がどのように制御されているのかという問いに重要な知見を与えるものです。
2. 研究の背景
胎児の初期発生において、シグナル伝達分子の濃度勾配が体軸を決定し、その後の組織形成を促します。細胞が受け取るシグナルの差異がどのように生じ、組織形成へとつながるのかを明らかにすることは、組織形成の基本原理を理解するうえで重要です。
TJP1などの細胞間接着分子は他の分子と複合体を形成し、タイトジャンクションを構築することで、上皮系組織の構造を保持しています。この複合体は、細胞内の足場タンパク質と膜貫通タンパク質によって構成され、上皮細胞どうしを結合させることで、層状の上皮組織を上側(頂端膜)と下側(基底外側膜)に区画化し、上皮細胞に上下の極性を形成し、高分子が細胞の隙間から反対側に自由拡散することを防いでいます。
原腸陥入の前段階であるエピブラストから原腸陥入への移行を制御するBMP4などの複数のシグナル伝達分子(モルフォゲン)は、タイトジャンクションで区切られた細胞の下部側(基底膜側)に分泌され、局在します。一方、BMP4の阻害因子であるNOGGINは上部側(頂端膜側)に分泌されます。
このことはエピブラストの上皮系構造と細胞極性が、モルフォゲンの濃度勾配の形成に重要な役割を果たし、さらに、モルフォゲンに対する非対称的な細胞反応を介した細胞分化のパターン形成に関与していることを示唆しています。また、過去の研究では、モルフォゲンが自発的に濃度勾配を形成し、それにより原腸陥入時のパターン形成が行われることが示唆されています。しかし、原腸陥入時の細胞間接着とモルフォゲンの濃度勾配との相互作用や複雑なパターン形成の具体的な制御メカニズムは未解明でした。
特定の培養条件のもと、ヒト多能性幹細胞のコロニーにBMP4を作用することで、原腸陥入でみられる細胞分化のパターン形成に似た状態を構築することができます。本研究は、この多能性幹細胞の分化誘導を、培養系での原腸陥入モデルとして用い、パターン形成における細胞接着の機能を解析しました。
その結果、細胞接着を制御するタンパク質複合体の構成因子の一つであるTJP1タンパク質の発現を特異的に抑制すると、多能性幹細胞のBMP4に対する反応性が亢進し、原腸陥入に類似した分化パターン形成が消失することがわかりました。このことは、ヒト多能性幹細胞による原腸陥入モデルで、TJP1が細胞分化のパターン形成を決定する重要な機能の一端を担っていることを示唆しています。
また、TJP1の発現を抑制した多能性幹細胞をBMP4処理して得られた分化細胞を詳細に解析したところ、始原生殖細胞に類似した細胞に分化していることがわかりました。この分化傾向は、TJP1の発現抑制によりBMP4シグナル伝達経路の活性化が長時間にわたり亢進したことに主に起因することが明らかになりました。
本研究の結果は、細胞間接着が胎児初期発生においてシグナル伝達および細胞分化と形態のパターン形成を積極的に制御している可能性、そして、ヒト多能性幹細胞の上皮構造が培養系での分化誘導法に大きく影響を与えることを示唆しています。
3. 研究結果
1)TJP1の発現抑制は、多能性幹細胞のBMP4に対する反応性を亢進させ、原腸陥入に類似した分化パターン形成を消失させる
はじめに、研究チームが以前に報告した、均質なコロニーに成長させたヒト多能性幹細胞をBMP4処理し原腸陥入に類似した分化パターンを誘導できるモデル(原腸陥入モデル)の再現性を確認しました(図1A)。
次に、抗生物質の一種であるドキシサイクリン(Doxycycline:Dox)の添加によってTJP1遺伝子を効率よく特異的に発現抑制できるヒトiPS細胞株(TJP1 CRISPRi iPS細胞株)を樹立しました(図1B)。Dox添加によってTJP1の発現を抑制したiPS細胞(TKD)の均質化コロニーをBMP4で処理したところ、TJ P1発現を抑制していない野生型iPS細胞(TWT)の均質化コロニーと比較して、iPS細胞のBMP4に対する活性化が長時間にわたり持続しました(図1C)。同時に、Dox添加によるTJP発現の抑制の結果、原腸陥入に類似した分化パターンが消失することがわかりました(図1D)。

図1 TJP1の発現抑制は、多能性幹細胞のBMP4に対する反応性を亢進させ、
原腸陥入に類似した分化パターン形成を消失させる
2)TJP1の発現抑制は、細胞間接着のバリアー機能を低下させることで多能性幹細胞のBMP4反応性を亢進させる
BMP4のシグナルを細胞内へ伝えるBMP4受容体は、上皮系構造の下部側、つまり上皮系細胞の基底膜側に局在します。そこで、培養細胞の上部と下部それぞれに作用できるトランスウエルメンブレン(図2A挿入図)を用いて、iPS細胞におけるBMP4受容体の局在します。そこで、過性の膜で、培養細胞の底面からも薬剤などを作用することのできるトランスウエルメンブレン(図2A挿入図)を用いて、iPS細胞におけるBMP4受容体の局在を確認しました。
野生型iPS細胞では、BMP4を下部側から作用した場合のみ、BMP4シグナル伝達の強い活性化が認められました。一方、TJP1の発現を抑制したiPS細胞ではどちらの側からBMP4を作用した場合も、強い活性化が長時間にわたり認められました(図2A)。このことから、TJP1の発現を抑制したことによって、①タイトジャンクションによる細胞膜の上側と下側の極性(区画化)が乱れてBMP4 受容体の局在が変化した可能性、あるいは、②細胞間接着が低下することにより、上部と下部の高分子の移動を制限するバリアー機能が低下し、上部に局在するシグナル分子が下部にある受容体に作用した可能性が考えられました。
まず、①を検証するために上部側マーカーを確認したところ、上部側マーカーの細胞内局在について、TJP1抑制による大きな影響はないことがわかりました。また、BMP4受容体自体を染色しても、TJP1抑制により、受容体の下部局在は変化しませんでした(図2B)。次に、②を検証するために上皮系構造の透過性を調べたところ、TJP1抑制により透過性が著しく増大することがわかりました(図2C, 2D)。
以上より、細胞極性の多少の乱れは考えられるものの、TJP1発現抑制によるBMP4シグナル伝達の増強、長期化は細胞のバリアー機能低下に起因していることが強く示唆されました。
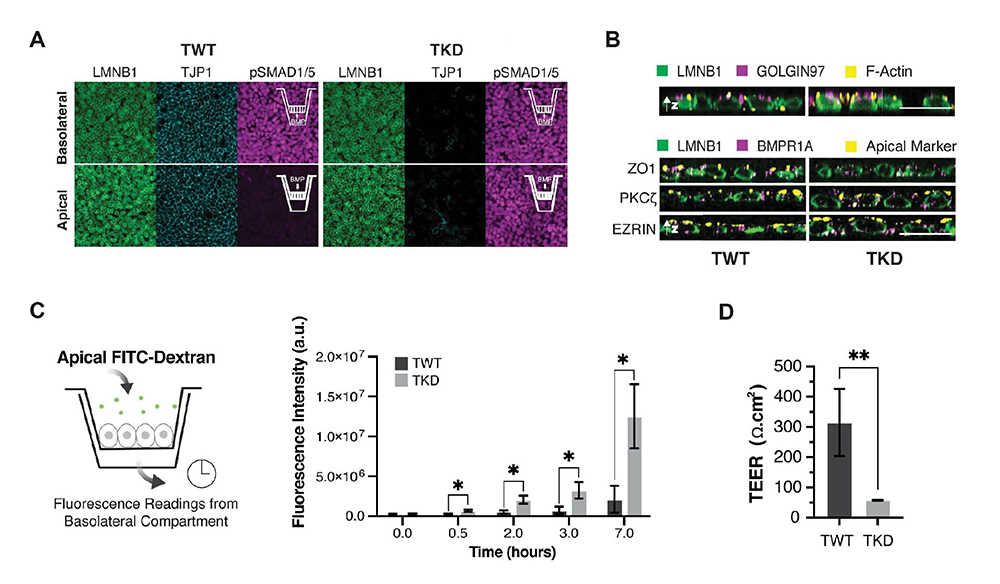
図2 TJP1の発現抑制は、細胞間接着のバリアー機能を低下させることで
多能性幹細胞のBMP反応性を亢進させる
3)TJP1の発現抑制は、多能性幹細胞を始原生殖細胞に類似した細胞に分化しやすくする
次に、TJP1抑制iPS細胞均質化コロニーをBMP4処理して得られた細胞群の網羅的遺伝子発現解析を行いました。その結果、TJP1発現抑制により、始原生殖細胞(PGC)への分化を制御する遺伝子群の発現が増加したことがわかりました(図3A)。さらに、BMP4処理によるPGCマーカーのタンパク質発現量を解析した結果、野生型iPS細胞均質化コロニーと比較して、TJP1抑制iPS細胞均質化コロニーでPGCマーカーの発現上昇がみられました(図3B)。
このPGC様細胞への分化傾向は、TJP1抑制iPS細胞均質化コロニーをBMP4で処理した場合のみでなく、通常の培養条件で培養したTJP1抑制iPS細胞をBMP4処理した条件下でも認められました(図3C)。以上のことから、接着機能を低下させることで、多能性幹細胞がPGC様細胞へと分化しやすくなる可能性が示唆されました。
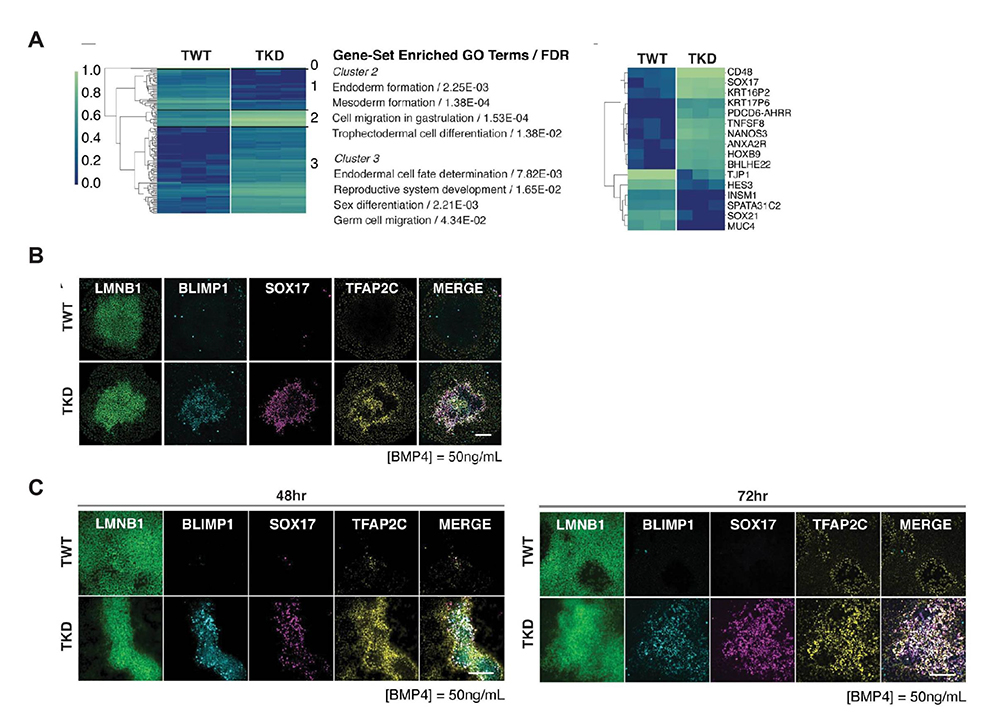
図3 TJP1の発現抑制は、多能性幹細胞を始原生殖細胞に類似した細胞に分化しやすくする
4)TJP1の発現抑制は、多能性幹細胞のBMP4への反応性を増強することにより、始原生殖細胞様細胞への分化を促進する
通常のヒト多能性幹細胞からPGC様細胞への分化は非常に効率が低く、その効率は1から2%にとどまります。多能性幹細胞からPGC様細胞へ効率よく分化誘導するためには、より未分化な状態に初期化した多能性幹細胞を使用する方法と分化誘導において初期中胚葉様細胞を経由する方法がこれまでに報告されています。
PGC様細胞への分化が促進される要因を詳細に理解するため、TJP1抑制にiPS細胞をより初期化させる効果があるかを解析しましたが、大きな影響は認められませんでした。また、TJP1抑制iPS細胞からPGC様細胞への分化過程で、初期中胚葉様細胞への分化を経由している形跡も認められませんでした。以上より、TJP1の発現抑制は、多能性幹細胞のBMP4シグナル伝達経路への反応性を増強することによって、PGC様細胞への分化を促進している可能性が考えられました。
この可能性をさらに検証するために、TJP1の発現抑制によらず、BMP4に対する反応性を増強させた場合も、PGC様細胞へと効率よく分化するかどうかを調べました。まず、トランスウエルメンブレン上で培養し、上下両側からBMP4を作用させたところ、野生型iPS細胞においても、BMP4シグナル伝達経路の強く長時間にわたる活性化が観察できました(図4A)。そして、この処理によって得られた分化細胞は、TJP1発現抑制iPS細胞をBMP4処理して得られた分化細胞と遺伝子発現パターンが非常に似ていることを確認しました(図4B)。
このことから、TJP1抑制による効果とトランスウエルメンブレンを用いたBMP4シグナル伝達活性化の効果がほぼ等しいと考えられ、TJP1抑制によるPGC様細胞への分化傾向がBMP4シグナル伝達の長時間にわたる強い活性化に起因している可能性が高まりました。
最後に、iPS細胞をトランスウエルメンブレン上で培養し、上下両側から低濃度のBMP4で処理すると、野生型iPS細胞でも効率よくPGC様細胞へと分化することがわかりました(図4C)。以上より、TJP1抑制により多能性幹細胞内でBMP4シグナル伝達経路が強く長時間活性することでPGC様細胞への分化が促進されていると考えられます。
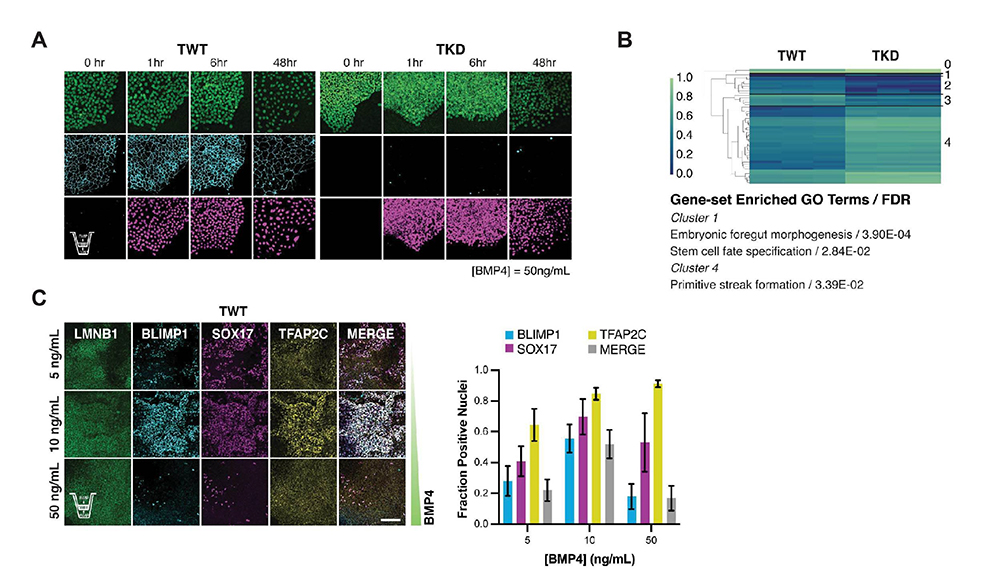
図4 TJP1の発現抑制は、多能性幹細胞のBMP4への反応性を増強することにより、
始原生殖細胞様細胞への分化を促進する
4. まとめ
本研究は、ヒト多能性幹細胞均質化コロニーから分化誘導する原腸陥入モデルを用いて、初期胚発生のパターン形成における上皮系構造と細胞接着の重要性を明らかにしました。また通常の多能性幹細胞から効率よく始原生殖細胞様細胞を作製できる可能性を示しました。これらの発見により、パターン形成メカニズムの理解がさらに進み、再生医療のためのヒト多能性幹細胞を用いた組織形成技術が発展することが期待されます。
5. 論文名と著者
- 論文名
Loss of TJP1 disrupts gastrulation patterning and increases differentiation toward the germ cell lineage in human pluripotent stem cells - ジャーナル名
Developmental Cell - 著者
Ivana Vasic1,2, Ashley R.G. Libby1,3, Annie Maslan2,4,5, Emily A. Bulger1,3, David Zalazar1,
Martina Z. Krakora Compagno1, Aaron Streets2,4,5,6, Kiichiro Tomoda1,7, Shinya Yamanaka1,7*,
Todd C. McDevitt1,8*
*: 責任著者 - 著者の所属機関
- グラッドストーン研究所 心血管研究部門
- UC Berkeley-UC San Francisco Graduate Program in Bioengineering, カリフォルニア大学
- Developmental and Stem Cell Biology Ph.D. Program, カリフォルニア大学
- Department of Bioengineering, カリフォルニア大学
- Center for Computational Biology, カリフォルニア大学
- チャン・ザッカーバーグ・バイオハブ
- 京都大学iPS細胞研究所(CiRA)
- Department of Bioengineering and Therapeutic Sciences, カリフォルニア大学
6. 本研究への支援
本研究は、下記機関より支援を受けて実施されました。
- 日本医療研究開発機構(AMED)再生医療実現拠点ネットワークプログラム「iPS細胞研究中核拠点」(JP21bm0104001)
- iPS細胞研究基金
- 京都大学On-site Laboratory
- 三木谷浩史
- Marc and Lynne Benioff
- L.K. Whittier Foundation
- Rodenberry Foundation
- The National Heart, Lung, and Blood Institute
- The National Institutes of Health
(U01-HL100406, U01-HL098179, R01-HL130533, and R01-HL135358) - The California Institute for Regenerative Medicine (LAI-C1408015)
- Emergent Behaviors of Integrated Cellular Systems consortium (CBET0939511)
7. 用語説明
注1)細胞間接着
隣りあう細胞どうしが接着すること。密着結合やギャップ結合など複数の様式がある。細胞接着分子が細胞間で結合することによって形成される。
注2)始原生殖細胞
卵と精子(配偶子)のもとになる細胞。英語では、primordial germ cell(PGC)という。
注3)原腸陥入
ヒトでは、受精のおよそ2週間後の初期胚で観察される細胞の分化と移動をともなう現象。原腸陥入により、細胞が外胚葉、中胚葉、内胚葉の三胚葉へと分化し、体軸が形成される。
注4)パターン形成イベント
細胞分化の非対称性や胚の形態を作りだす初期胚発生の現象。原腸陥入のほかに、体節形成などがある。
注5)タイトジャンクション(密着結合)
血管や腸管などの上皮組織でみられる細胞間接着の様式。上皮系細胞が層状に配置された上皮組織の外側(頂端膜)と内側(基底外側膜)を規定し、内外を隔てることで高分子の往来を制限するバリアーとしての機能を持つ。
注6)TJP1
タイトジャンクションを形成する分子複合体を構成する細胞質タンパク質の一つ。他の細胞との結合をになう細胞膜タンパク質や細胞骨格などと結合する。TJP1はTight junction protein-1の略。ZO1(Zonula occludens-1)ともよばれる。

