2019-11-22 基礎生物学研究所,理化学研究所
窒素は全ての生物が生命を維持するために必須な成分です。一般に植物は硝酸塩やアンモニアといった窒素栄養素を土壌から吸収します。一方、マメ科植物は、根粒と呼ばれる特殊な器官に窒素固定細菌を共生させており、ほとんどの生物が利用できない空気中の窒素を栄養素として使うことができます。そのために、マメ科植物は窒素栄養素が乏しい痩せた土地でも生育することができます。 根粒共生は、植物にとっても、またその恩恵に預かる我々にとっても大変有用な形質ですが、マメ科植物とマメ科に近縁な一部の植物だけで見られる現象です。これまでの研究により、根粒共生に関わる遺伝子についての情報は次第に蓄積されてきていますが、マメ科植物の根粒共生の能力が進化の過程でどのように獲得されてきたのかは、よく分かっていませんした。基礎生物学研究所 共生システム研究部門の征矢野敬准教授と理化学研究所 環境資源科学研究センター 植物共生研究チームの林誠チームリーダーは、基礎生物学研究所 川口正代司教授、農業・食品産業技術総合研究機構 下田宜司主任研究員と共同で、根粒共生における根粒の形成過程に、植物が一般的に持つ側根の形成メカニズムの一部が流用されていることを新たに発見しました。マメ科植物モデル植物のミヤコグサを用いて、側根の形成を制御する遺伝子として知られるASL18a遺伝子が、他の根粒形成遺伝子と協調しながら根粒の発達を制御することを明らかにしました。これは、マメ科植物の根粒共生の能力が、植物に一般的な既存のシステムを上手く取り入れながら獲得されてきたことを示すものです。この成果は、2019年11月22日に米国の科学雑誌『Science』に掲載されます。

成果の概念図:根粒形成と側根の形成は共通の遺伝子ALS18aによって制御される
【研究の背景】
我々が植物を栽培する時には、植物が元気に育つように肥料を与えます。肥料には硝酸塩やアンモニアといった窒素栄養素が含まれています。多くの植物は側根と呼ばれる枝根を発達させることで、土壌中の限られた窒素栄養素を効率的に吸収できるように工夫しています。一方、マメ科植物は窒素固定細菌と共生することで、ほとんどの生物が利用できない空気中の窒素を栄養素として直接的に使うことができます。そのためにマメ科植物は他の植物では成長が抑制されるような窒素栄養の乏しい土壌でもすくすくと育ちます。
マメ科植物と共生する窒素固定細菌は根粒菌と呼ばれ、根粒菌が宿主の根に感染すると根粒と呼ばれる瘤が根に形成されます。根粒は共生器官であり、根粒菌を宿主の細胞に取り込むことで、根粒菌が窒素固定するための丁度良い環境を提供します。したがって、根粒は根粒共生の要となる特別な器官と言えます。根粒共生はマメ科植物とマメ科植物に系統的に近縁な極限られた植物のみに見られることから、マメ科植物が進化の過程で根粒を形成する仕組みをどのように発達させてきたのかが長い間の疑問でした。
【研究の成果】
研究グループは、これまでの研究において、ミヤコグサの根粒形成にNIN転写因子と呼ばれる遺伝子の機能が重要であることを明らかにしていました。しかし、NINが具体的にどのように作用しているのかが分かっていませんでした。そこで今回、NIN転写因子によって働きが調節される遺伝子を探索しました。その結果、NINの下流で、ASL18aとNF-Yという性質の異なる2つの転写因子が協調的に作用して根粒の形成を制御することを新たに発見しました(概念図)。
NF-Yは核を持つ全ての生物が持っており、様々な生命現象に関係していることが知られている転写因子です。NF—Yが失われたミヤコグサ変異体(nf-ya1 nf-yb1変異体)では、根粒の発達が抑制されました(図1)。
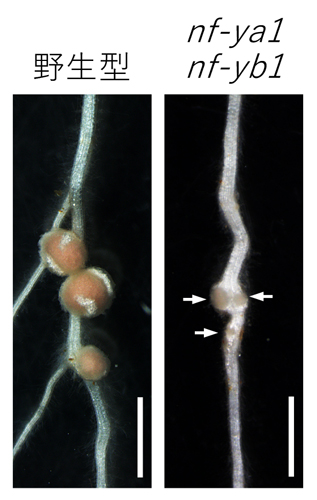
図1:NF—Yの根粒形成における機能
根粒菌接種15日目の野生型(左)、nf-ya1 nf-yb1二重変異体(右)。矢印は未熟な根粒を示す。スケールバー: 1 mm。
NF—Yが失われたミヤコグサ変異体(nf-ya1 nf-yb1二重変異体)では、根粒の発達が抑制される。
ASL18aは、シロイヌナズナやイネなどの非マメ科植物では側根や冠根を作るときに働くことが知られています。ミヤコグサにおいてその働きを調べると、ASL18aは側根に加えて根粒ができるときにも働いていることが分かりました(図2)。ASL18aを失ったミヤコグサ変異体(asl18a変異体)では、側根の発達が抑制されました(図3)。NF—YとASL18aの両方を失った変異体では、NF—Yだけを失った変異体(nf-ya1 nf-yb1変異体)で見られる根粒の発達抑制がより重篤になりました(図4)。
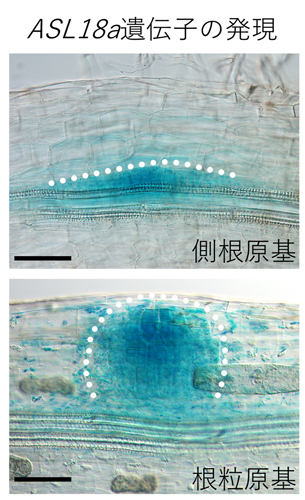
図2:ASL18a遺伝子は側根形成時と根粒形成時に誘導される
ASL18aが発現する細胞を青色に染色した。未熟な側根と根粒(側根原基と根粒原基)の位置を破線で示している。スケールバー:0.1 mm

図3:ASL18aの側根における機能
播種14日目の野生型の根(左)とasl18a変異体の根(右)。スケールバー:1 cm。asl18a変異体では側根形成が抑制される。
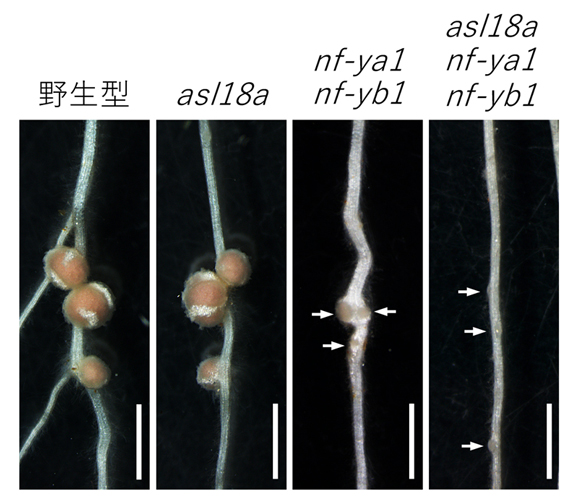
図4:ASL18aとNF—Yの根粒形成における機能
左から根粒菌接種15日目の野生型、asl18a変異体、nf-ya1 変異体、nf-ya1 nf-yb1二重変異体asl18a nf-ya1 nf-yb1三重変異体の根粒。矢印は未熟な根粒を示す。スケールバー: 1 mm。
NF—Yが失われたミヤコグサ変異体(nf-ya1 nf-yb1二重変異体)では、根粒の発達が抑制される。NF—YとASL18aの両方を失った変異体(asl18a nf-ya1 nf-yb1三重変異体)では、NF—Yだけを失った変異体(nf-ya1 nf-yb1二重変異体)で見られる根粒の発達抑制がより重篤になった。
さらに研究グループは、NF-YとASL18aは直接結合する性質を持つことを明らかにしました。そして、根粒ができないはずのnin変異体において、ASL18aとNF-Yをミヤコグサの根で同時に働かせることで、根粒を付けさせることに成功しました(図5)。このことは、ASL18aとNF-Yが一緒に働くことでNINの代わりをしたことを意味します。
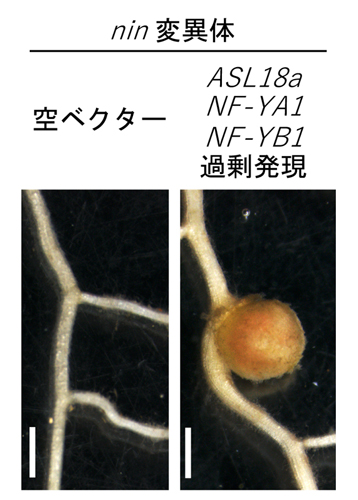
図5:ASL18aとNF-Yによるnin変異体の根粒形成不全の回復
nin変異体では根粒菌がいても根粒は形成されない(左)。根粒ができないnin変異体に、ASL18aとNF-Yを強制的に発現させると、根粒形成能が回復し、根粒が形成された(右)。スケールバー:1 mm。
また、根粒ができるときにNINがASL18a遺伝子の特定の場所に結合することでASL18a遺伝子が働き始めることも分かりました。NINが結合する特定の場所は、ダイズやインゲンなどのミヤコグサ以外のマメ科植物のASL18a遺伝子にも共通して見つかりました(図6)。しかし、非マメ科植物のASL18a遺伝子では見つかりませんでした。このNINが結合する場所をマメ科植物が進化の過程で獲得したことが、ASL18の根粒での機能に影響したことが示唆されます。
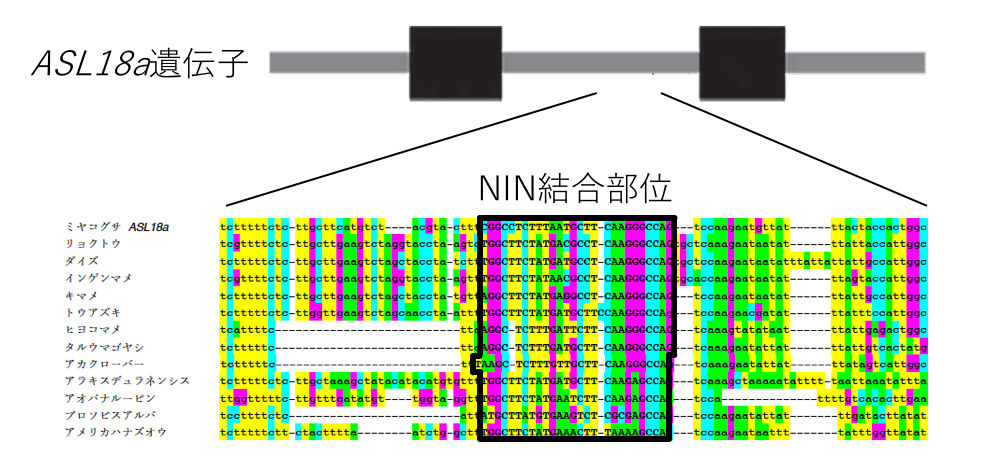 図6:ASL18a遺伝子の構造とマメ科植物のASL18遺伝子にあるNIN結合DNA配列
図6:ASL18a遺伝子の構造とマメ科植物のASL18遺伝子にあるNIN結合DNA配列
ASL18遺伝子にあるNIN結合配列はマメ科植物に共通して存在する一方で、非マメ科植物には見られない。
以上のことから、本来は側根の形成を制御するASL18a遺伝子が、根粒菌が根に感染するとNINの指令により働きはじめ、ASL18aとNF-Yとが相互作用しながら根粒の発達を制御することが分かりました(図7)。
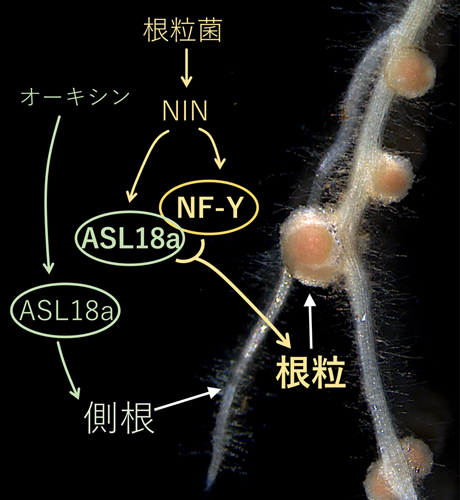
図7(成果の概念図):根粒形成と側根の形成は共通の遺伝子ALS18aによって制御される
【今後の展望】
近年の研究から、マメ科植物は他の植物でも共通に持っている因子を上手に使いながら根粒共生を制御していることが少しずつ分かってきています。今回の成果は、側根の形成メカニズムの一部がASL18aを介して根粒共生に流用されていることを示しています。根粒形成の基本的な分子機構は特別ではなく、マメ以外の植物とさほど大きな違いはないのかもしれません。さらに研究を掘り下げることで、根粒共生の進化の道筋のさらなる理解や非マメ科植物に根粒を付けさせるような挑戦的な研究に発展すると期待されます。
【発表雑誌】
雑誌名: Science
掲載日: 2019年11月22日
論文タイトル: A shared gene drives lateral root development and root nodule symbiosis pathways in Lotus
著者: Takashi Soyano, Yoshikazu Shimoda, Masayoshi Kawaguchi, Makoto Hayashi
DOI: 10.1126/science.aax2153 (https://science.sciencemag.org/content/366/6468/1021)
【研究グループ】
本研究は、基礎生物学研究所 共生システム研究部門の征矢野敬准教授、川口正代司教授、理化学研究所 環境資源科学センター 植物共生研究チームの林誠チームリーダー、農業・食品産業技術総合研究機構 生物機能利用研究部門の下田宜司主任研究員の共同研究として実施されました。
【研究サポート】
本研究は科学研究費助成事業 (16K08149) 、理化学研究所奨励課題、日本学術振興会 最先端・次世代研究開発支援プログラム (GS029)のサポートを受けて実施されました。また、基礎生物学研究所 生物機能解析センター、モデル生物研究センターの協力を得ながら実施されました。
【本研究に関するお問い合わせ先】
基礎生物学研究所 共生システム研究部門
准教授 征矢野 敬(そやの たかし)
理化学研究所 環境資源科学センター 植物共生研究チーム
チームリーダー 林 誠(はやし まこと)
【報道担当】
基礎生物学研究所 広報室
理化学研究所 広報室 報道担当

