
2019-11-28 東京大学
島津 舜治(生物科学専攻 修士課程1年生)
瀬尾 光範(理化学研究所 環境資源科学研究センター ユニットリーダー)
寺島 一郎(生物科学専攻 教授)
矢守 航(現:大学院農学生命科学研究科 生産・環境生物学専攻 准教授
/研究当時:生物科学専攻 准教授)
発表のポイント
- 光合成能力を最大限に発揮するまでのウォーミングアップにあたる“光合成誘導”の時間を比較したところ、全身に光を浴びる植物個体では、葉一枚のみに光が当たる個体に比べ、所要時間が短縮されることを発見した。
- 個体内で光の当たる葉の枚数が増加するにつれて、光の当たった葉の通気口である気孔がすばやく開き、光合成誘導の時間が短縮された。変異体を用いた解析から、光の当たる葉の枚数の増加にともない、葉一枚あたりの気孔閉鎖ホルモン・アブシシン酸の量が減少することが示唆された。
- 本研究成果は、個体の中でも一枚の葉のみを対象とした従来の光合成研究に対する警鐘となるとともに、植物工場などにおける光照射法の最適化にも貢献する。
発表概要
太陽が昇ると、植物の1日もスタートする。暗闇にいた植物は光を受けると同時に光合成を開始するが、光合成能力を最大限に発揮するまで植物はウォーミングアップを必要とする。この期間は“光合成誘導(注1)”とよばれ、その間、植物は吸収した光エネルギーを最大限有効利用できない。ヒトが朝起きてすぐには行動を開始できないように、光合成誘導は植物にとっての“寝起きでうまく動けない時間”と言える。太陽が昇る早朝のみならず、日中でも畑や林床などでは、雲の切れ間から降り注ぐ光や、風で揺らめく植物の葉の間から差し込む光の影響で、植物の受ける光量は頻繁に変動しており、一日の中でも光合成が弱光下のオフの状態から強光下のオンの状態になる誘導現象が見られる。したがって、野外の変動する光環境において光合成誘導期間を短縮することによって植物生産性を向上させようという取り組みが世界中でなされている。
東京大学大学院理学系研究科の島津舜治 大学院生(研究当時:学部生)、大学院農学生命科学研究科の矢守航 准教授(研究当時:大学院理学系研究科 准教授)らは、複数の植物種を用いて、全身に光を受けた植物個体では、葉一枚のみに光を受けた個体に比べて光合成誘導期間が短縮されることを見出した(図1)。

図1:全身に光を浴びることが植物の光合成効率を改善する
さらに、光合成誘導の期間の短縮は、葉にCO2を取り込む可動式の通気口である気孔(注2)が光に応じてより俊敏に開口し、葉内のCO2不足がすばやく解消されることによって起こることを突き止めた。また、シロイヌナズナの各種変異体を用いた解析から、気孔開閉に関わる植物ホルモンABA(注3)がこの現象に関与していることを明らかにした。
本研究では、同一個体内でも光の当たり具合で植物の“寝起き”が良くなることを明らかにした。これまでの葉の光合成に関する研究では、一枚の葉(あるいはその一部)を透明の箱に密閉して光合成誘導を含む光合成の諸性質が解析されてきた。今回の研究は、光合成誘導を植物個体全体の現象として初めて捉えたものであり、“野外での植物の自然な振る舞い”の理解に一歩近づくものである。また、本研究で得られた知見は、植物工場などにおける光照射法の最適化にも貢献する。
発表内容
光合成は植物の成長量や収量を決定する最も重要な代謝であるため、太陽の光エネルギーを最大限に活用することは、植物の生存戦略上の至上課題であるといえる。暗黒下や弱光下に置いた植物に急に強光を照射すると、光合成速度は徐々に上昇し、やがて定常状態に達する(この現象を光合成誘導と呼ぶ)。暗黒下や弱光下では逆に光合成系が不活性化される。野外に生育する農作物や林床植物は、雲や植生によって光が遮られる結果、常に変動する光環境下に晒されており、頻繁に光合成系のスィッチオフと誘導とが起こる。また、光は植物全体に均一に照射されるのではなく、同一の植物個体内にも、光があたっている葉とあたっていない葉が見られるため、光のわずかな移り変わりによっても光合成誘導が起こる。光合成誘導の期間、植物は吸収した光エネルギーを有効利用できない。光合成が定常状態に達するまでの時間を短縮することができれば、変動する光環境において、吸収した光エネルギーを有効利用でき、植物生産性の向上に大きく貢献すると考えられる。
実験室で行う光合成研究は一枚の葉を対象として行われることが多く、同一個体内の他の葉の環境なども考慮した植物個体全体としての現象として光合成誘導を捉えることは稀であった。そこで本研究では、植物体全体への光照射と葉一枚への光照射に対する光合成誘導の違いを解析した。光合成活性をイメージ画像で捉えることのできる光合成可視化装置(Imaging-PAM)および高性能のガス交換光合成測定装置(Li-6400XT)を用いて、モデル植物のシロイヌナズナおよびイネにおいて、6時間以上暗処理した植物個体の光合成誘導を解析した。通常の測定条件である測定部位のみに照射した”IIL”条件(Individually Irradiated Leaf, 図2左)に加え、測定時に一定強度の光を植物体全体に照射した”WIP”条件(Whole Irradiated Plant, 図2右)の2条件で光合成誘導を比較した。
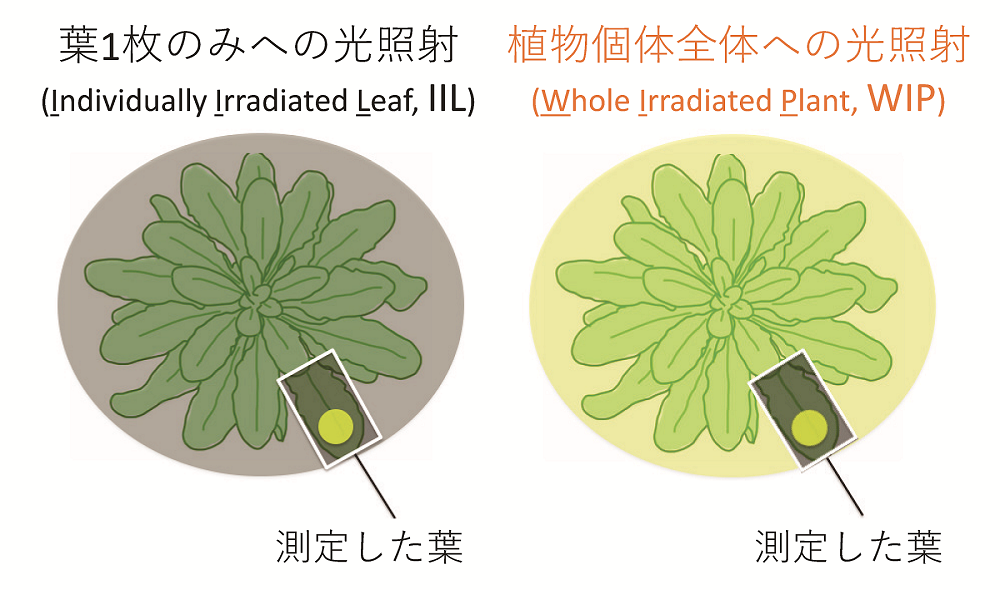
図2:葉1枚のみへの光照射(IIL条件, 左)と植物個体全体への光照射(WIP条件, 右)
光合成測定を行う特定の一枚の葉は均一な環境に設定し、測定部位以外の周辺環境を変化させた。測定部位にのみ光を照射した一般的な測定方法である”IIL”条件に対し、測定部位に加えて同強度の光を植物体全体に照射した”WIP”条件の2条件で光合成誘導を比較した。
その結果、通常の葉一枚のみを照射したIIL条件に比べ、個体全体を照射したWIP条件において、光合成誘導の短縮が見られることを光合成可視化装置で確認し(図3)、高性能の光合成解析装置によりさらに定量的に解明することができた(図4A)。また、個体全体照射によって光合成誘導が短縮された原因は、葉にCO2を取り込む可動式の通気口である気孔が光に応じてより俊敏に開口し、葉内のCO2不足が素早く解消されるためであることを突き止めた(図4B)。
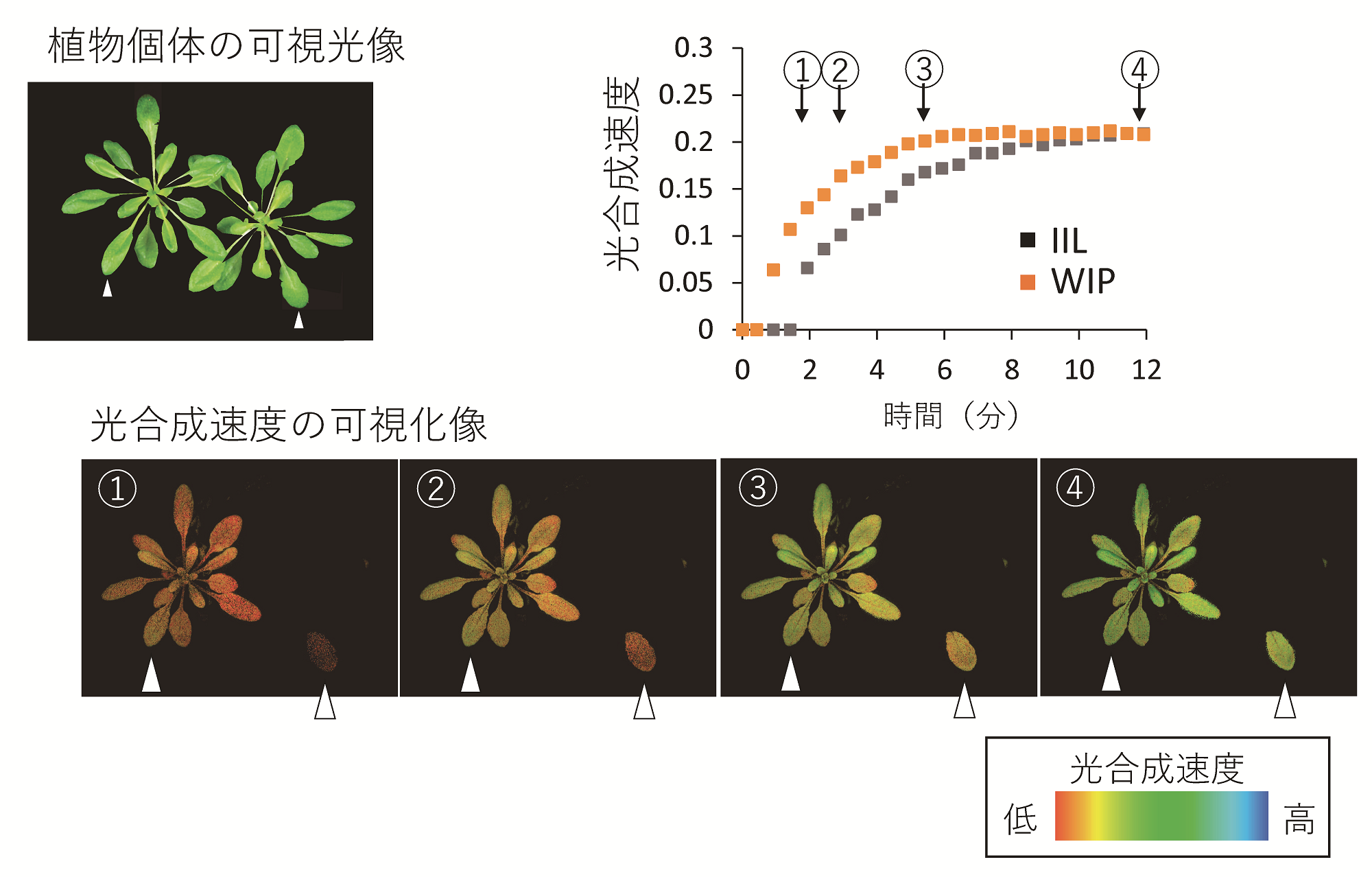
図3:光合成反応の可視化によって捉えられた、植物個体全体への光照射(WIP条件)による光合成誘導の促進
光合成活性をイメージ画像で捉えることのできる光合成可視化装置(Imaging-PAM)を用いて、葉一枚のみ光照射したIIL条件(可視光像右)と、植物個体全体を照射したWIP条件(同左)における光合成誘導の違いを解析した。解析の対象として同程度生育した2個体の同じ葉位の葉(白矢尻)を用い、光合成活性の指標の一つであるY(II)を計測したところ、WIP条件でより素早く低活性状態(オレンジ色)を脱し、定常状態(緑色)に至る様子が捉えられ、各タイムポイントにおける測定値の差が如実に可視化される結果となった。
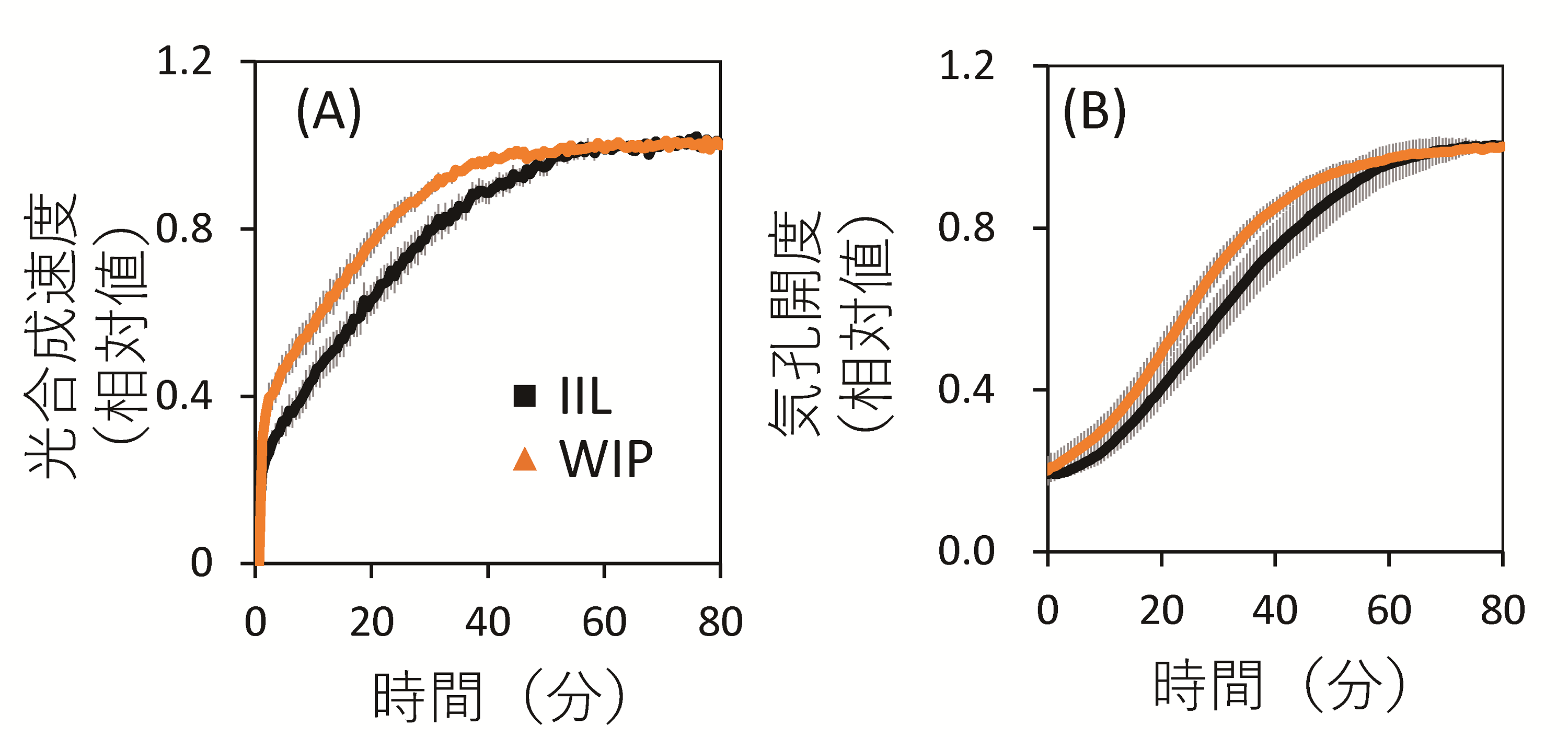
図4:植物個体全体への光照射(WIP条件)によって、光合成誘導と気孔開口は促進される
ガス交換光合成測定装置(Li-6400XT)を用いて、葉一枚のみ光照射したIIL条件と、植物個体全体を照射したWIP条件における光合成速度(A)および気孔開度(B)を解析した。気孔開度は気孔の開き具合を表す数値で、大きければそれだけ気孔が開いていることを示す。両指標とも定常状態の値を1とした相対値を算出し比較したところ上図のように、定常状態に至るまでに要する時間に大きな違いを示した。定常状態の60%の値に至るまでの所要時間を算出し統計的な解析を行なったところ、両指標で有意な差が見られた。
さらなるメカニズム探究のため、気孔に作用する植物ホルモンであるABAトランスポーター(注4)の欠損変異体であるabcg25とabcg40(注5)を用いて同様の実験を行なったところ、葉一枚のみを照射したIIL条件と個体全体を照射したWIP条件で、光合成誘導と気孔開口速度に違いは見られなかった(図5)。これらの結果は、個体全体照射による光合成誘導の促進に、ABAが関与していることを示す。
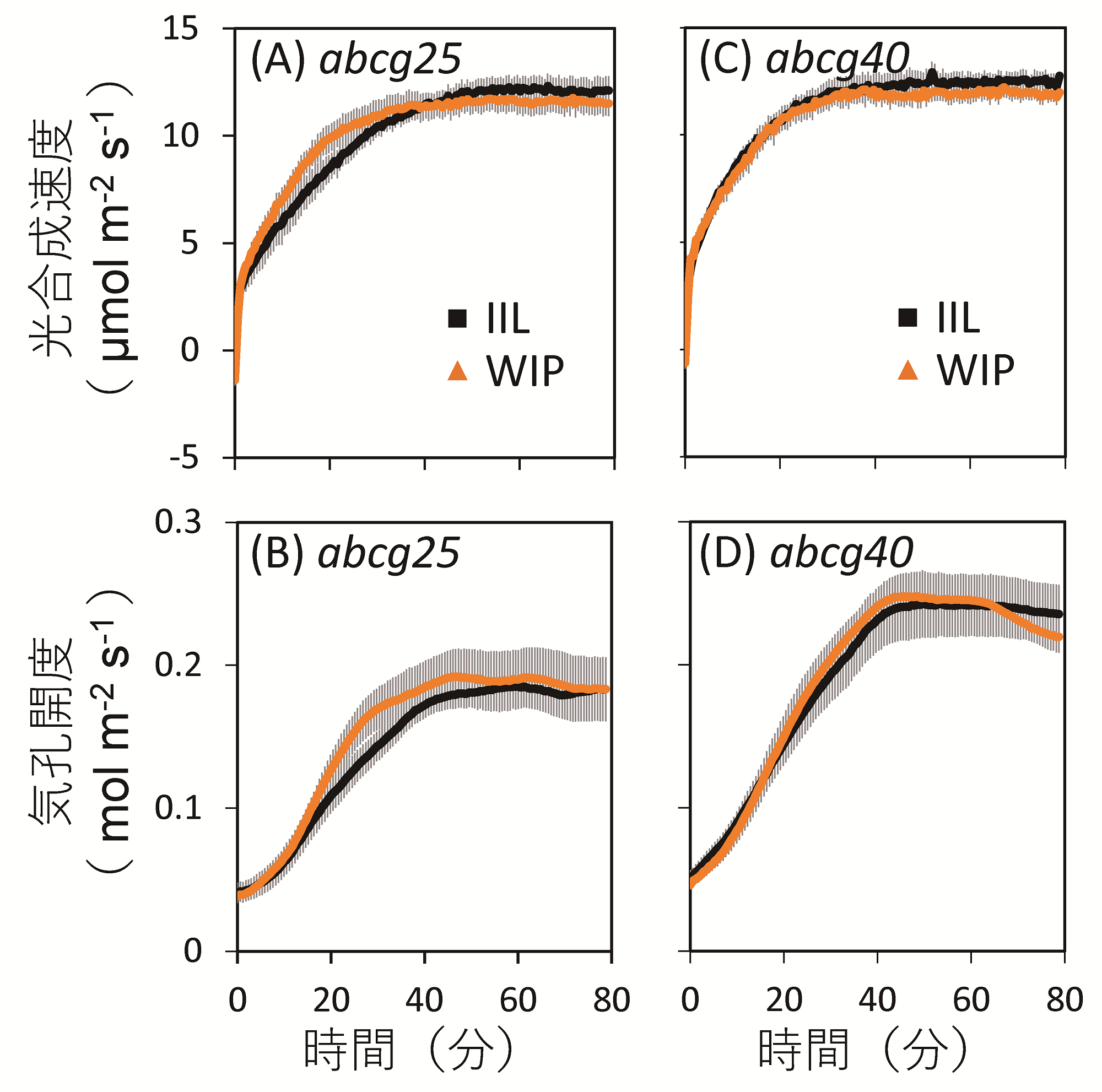
図5:abcg変異体における植物個体全体への光照射(WIP条件)が光合成誘導に及ぼす影響
ABAトランスポーターの欠損変異体であるabcg25とabcg40変異体において、ガス交換光合成測定装置(Li-6400XT)を用いて、葉一枚のみ光照射したIIL条件と、植物個体全体を照射したWIP条件における光合成速度および気孔開度を測定した。その結果、両変異体ともに、IIL条件とWIP条件とでは、光合成誘導(A, C)および気孔開口(B, D)に大きな違いは見られなかった。定常状態の60%の値に至るまでの所要時間を算出し統計的な解析を行なっても同様に有意な差は検出されなかった。
本研究を通して、植物は光の当たり具合に応じて個体内で情報伝達を行い、光合成誘導の速さを変えることが示唆された。今回の成果は、葉の光合成を研究する際には、一枚の葉の環境のみではなく個体全体の環境を考慮する必要があることも示している。野外の植物の自然な振る舞いをより詳細に理解するためにも、また、成長量の増大に直結する光合成誘導を改善するためにも、植物の“光照射法”の重要性が認識されなければならない。
発表雑誌
雑誌名
Frontiers in Plant Science論文タイトル
Whole irradiated plant leaves showed faster photosynthetic induction than individually irradiated leaves via improved stomatal opening著者
Shunji Shimadzu, Mitsunori Seo, Ichiro Terashima and Wataru Yamori*DOI番号
10.3389/fpls.2019.01512
アブストラクトURL
https://www.frontiersin.org/articles/10.3389/fpls.2019.01512/abstract
用語解説
注1 光合成誘導
光合成が始動し、定常値に達するまでのCO2固定速度の変化。この期間は光エネルギーを有効利用することが出来ない。この誘導反応が起こる理由として、光照射後に、光合成に関わるRubiscoなどの酵素群の活性化が順次起こることや、暗黒や弱光下で閉じていた気孔が徐々に開き、光合成反応の基質であるCO2の供給量が増えていくことなどが挙げられる。
注2 気孔
葉の表皮に主に存在する開閉式の小さい孔。周囲に位置する孔辺細胞という一対の細胞が膨張収縮することによって開閉し、植物体と大気間のガス交換(CO2吸収や蒸散)を可能にしている。一般に夜間は不要な蒸散を防ぐべく閉口し、光照射とともに開口していく性質がある。
注3 ABA
植物ホルモンの一つで、正式名称はアブシジン酸。ストレスホルモンと呼ばれ、気孔の開閉のほか、乾燥・低温などのストレス応答、種子の登熟などに機能する。主に維管束組織において、カロテノイドの酸化的解裂を経由して合成される。
注4 トランスポーター
細胞膜を横切って物質の輸送を媒介する内在性膜タンパク質で、それぞれ特定の物質を輸送する。糖やアミノ酸といった親水性物質は、疎水性の細胞膜を透過できないため、何らかのエネルギーを利用したトランスポーターによる輸送が行われる。
注5 abcg25 および abcg40
ABAトランスポーターの欠損変異体。ABCG25は維管束組織に位置し、ABAを組織外に排出する機能を持ち、ABCG40は孔辺細胞に位置し、ABAを取り込む機能を持つ。ABAは維管束組織で主に合成されるため、どちらの変異体もABAの孔辺細胞への輸送に異常をきたす。
―東京大学大学院理学系研究科・理学部 広報室―

