
2020-10-27 理化学研究所,鳥取大学
理化学研究所(理研)環境資源科学研究センターケミカルバイオロジー研究グループの尹忠銖研究員、本山高幸専任研究員、長田裕之グループディレクターと鳥取大学工学部化学バイオ系学科の永野真吾教授らの共同研究チームは、かび毒[1]として知られる「テヌアゾン酸[2]」の生合成酵素である「TAS1」の「ケト合成酵素(KS)ドメイン[3]」の立体構造を明らかにし、テヌアゾン酸生合成における鍵となる反応のメカニズムに関する重要な知見を得ました。
本研究成果は、テヌアゾン酸の高効率な生産制御を可能にすると期待できます。また、テヌアゾン酸の構造を変えることによる、有用な新しい生理活性物質の創出に繋がる可能性があります。
かび毒は植物病原糸状菌[4]などが生産する化学物質で、農産物汚染による健康被害や植物病害を引き起こすため、経済的損失が大きく問題となっています。
今回、共同研究チームは、イネいもち病菌[5]が持つかび毒テヌアゾン酸の生合成酵素TAS1のKSドメインの立体構造をX線結晶構造解析[6]により明らかにし、立体構造に基づく酵素の機能解析により、生合成の際の環化反応メカニズムに関する重要な知見を得ました。TAS1のKSドメインは、通常のKSドメインと同様のアミノ酸残基がユニークな反応に関与していること、また、通常のKSドメインの場合よりも大きな基質を受け入れるために進化していることが示唆されました。
本研究は、科学雑誌『Journal of Biological Chemistry』(8月14日号)に掲載されました。

イネいもち病菌がかび毒テヌアゾン酸を作るメカニズム
背景
「テヌアゾン酸」は、かび毒として知られる化合物(二次代謝産物[7])で、植物病原糸状菌の一種であるAlternaria(アルタナリア)[8]から単離・同定され、1957年に初めて報告されました。Alternariaは世界中に広く分布しており、さまざまな穀物や果物などに被害を及ぼしています。
テヌアゾン酸はAlternariaが感染した植物から検出されてきましたが、現在では、イネいもち病菌など多くの植物病原糸状菌がテヌアゾン酸を生産することが分かっています。テヌアゾン酸は、植物のタンパク質合成を阻害することで毒として作用します。テヌアゾン酸などのかび毒を制御するには、その生産メカニズムを明らかにすることが重要です。
これまで、尹忠銖研究員、本山高幸専任研究員、長田裕之グループディレクターらはイネいもち病菌を用いて、二次代謝活性化に関する研究を推進してきました。2015年には、テヌアゾン酸の生産誘導を引き起こし、発現誘導される遺伝子の中から生合成遺伝子TAS1を同定し、生合成メカニズムの概要を明らかにしました注1)。また2017年には、テヌアゾン酸の生産誘導に関与するタンパク質を同定することに成功しています注2)。
今回、共同研究チームは、テヌアゾン酸の生合成酵素「TAS1(tenuazonic acid synthetase 1)」の環化反応に必要とされる「ケト合成酵素(KS)ドメイン」の立体構造を明らかにし、テヌアゾン酸生合成における鍵となる反応のメカニズムを解明することを試みました。
注1)2015年10月27日プレスリリース「かび毒テヌアゾン酸の生合成遺伝子を同定」
注2)2017年10月6日プレスリリース「かび毒の生産制御機構」
研究手法と成果
多くの微生物の二次代謝産物は、ポリケチド合成酵素(PKS)[9]や非リボソーマルペプチド合成酵素(NRPS)[10]、およびそれらの融合酵素により作られます。TAS1は、糸状菌からは初めて見いだされたNRPS-PKS融合酵素であり、NRPS部分はC/A/PCPの三つのドメインから、PKS部分はKSドメインのみで構成されています。PKSのKSドメインは、通常はポリケタイド鎖の伸長反応を触媒します。一方、TAS1のKSドメインは、アミノ酸を含むイソロイシン・ジケチドを基質として、基質伸長反応を触媒せずに環化反応のみを触媒します(図1)。本研究では、その反応メカニズムを明らかにするため、TAS1のKSドメインの立体構造解析を行いました。
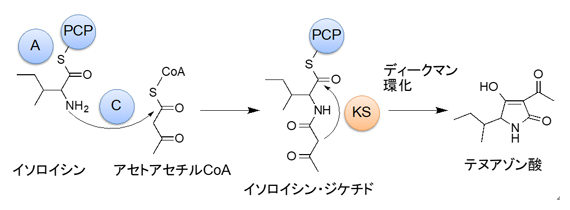
図1 テヌアゾン酸の生合成経路
イソロイシンとアセトアセチルCoAから、テヌアゾン酸生合成酵素TAS1によりテヌアゾン酸が作られる。イソロイシンとアセトアセチルCoAからC、A、およびPCPドメインを用いて生合成中間体(イソロイシン・ジケチド)が作られ、TAS1のKSドメインがテヌアゾン酸生合成の最終段階であるディークマン環化反応を行う。
まず、TAS1のKSドメインタンパク質を、大腸菌を宿主として異種発現し、精製することにより取得しました。現在までの研究で、PKSのKSドメインは二量体として機能していることが報告されていますが、ゲルろ過[11]による解析の結果から、TAS1のKSドメインは溶液中で単量体として存在することが明らかになりました。
次に、X線結晶構造解析により、1.68オングストローム(å、1åは100億分の1メートル)の分解能で立体構造を決定しました。決定した全体構造はI型PKS[12]のKSドメインと類似していましたが、基質ポケット(基質が結合する部位)の構造が異なっていました。一般的なI型PKSのKSドメインは、α-ヘリックス[13]が基質ポケットの上部を覆うように存在しているのに対し、TAS1のKSドメインにはこのα-ヘリックスがなく、そのために基質ポケットが大きく広がっていることが明らかになりました。類似の構造は、他のアミノ酸を含む化合物を基質とするオキサゾロマイシン[14]生合成酵素(OzmQ)のKSドメインでも報告されていることから、より大きな基質を受け入れるKSドメインに共通の特徴である可能性があります。
TAS1のKSドメインの活性残基として予測された179番目のシステイン(Cys179)、322番目のヒスチジン(His322)、376番目のアスパラギン(Asn376)、I型KSドメインとは異なる特異的残基である324番目のセリン(Ser324)、378番目のグルタミン酸(Glu378)の変異体解析と、基質であるイソロイシン・ジケチドをリガンドとしたドッキングシミュレーション[15]の結果から、リガンド結合部位の近傍に存在するHis322がプロトン(水素イオン)を引き抜くことで環化反応が誘起されることが示唆されました(図2)。
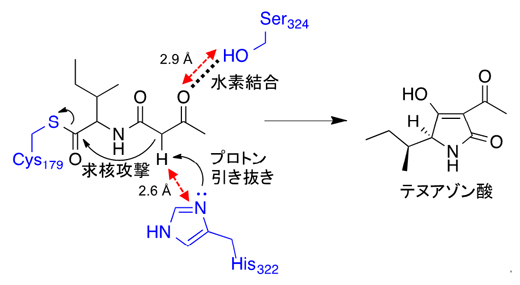
図2 TAS1のKSドメインによる環化反応のメカニズム
TAS1のKSドメインの結晶構造とドッキングシミュレーションの結果から、Cys179に結合した基質がSer324との水素結合により適切な方向に配置され、その後、His322によるプロトン(水素イオン)の引き抜きがチオエステルのカルボニルに対する求核攻撃を引き起こし、結果として環化とテヌアゾン酸の酵素からの放出を引き起こすことが示唆された。
さらに、さまざまな基質を用いた解析により、TAS1のKSドメインは通常のKSドメインと比較してより広範囲の基質を受け入れることができる特徴を持つことを見いだしました。この特徴は、酵素の基質ポケットの中の単一アミノ酸変異(378番目のグルタミン酸をアラニンに変更)により増強されることが明らかになりました。
本研究では、TAS1のKSドメインのユニークな特徴を見いだしました。以上の結果は、かび毒テヌアゾン酸の生合成においてKSドメインが行う環化反応のメカニズムをより深く知るためのヒントになります。また、NRPS-PKS融合酵素において、アミノ酸を含む基質を受け入れるKSドメインの機能を深く知るための手がかりとなります。
今後の期待
本研究の成果により、かび毒であるテヌアゾン酸の効率的な生産制御が可能になることが期待できます。例えば、生合成酵素TAS1に対する阻害剤を開発することにより、テヌアゾン酸の生産を抑制することが可能になります。
また、テヌアゾン酸はさまざまな生物に対して生理活性を示します。本研究の成果を応用することにより、テヌアゾン酸の構造を改変し、有用な生理活性物質を創出できる可能性があります。
補足説明
1.かび毒
マイコトキシンとも呼ばれ、糸状菌の二次代謝産物として生産される毒の総称。アフラトキシン、トリコテセンなどが知られ、農産物の汚染による健康被害や植物病害を引き起こすため経済的損失が大きく問題となっている。
2.テヌアゾン酸
かび毒として知られる二次代謝産物で、植物病原糸状菌Alternaria(アルタナリア)から単離・同定され、1957年に初めて報告された。Alternaria以外のさまざまな植物病原糸状菌もテヌアゾン酸を生産する。欧州食品安全機関(EFSA)はテヌアゾン酸に対する毒性学的懸念の閾値として一日あたり1,500ng/kgという値を示している。1ngは、10億分の1g。
3.ケト合成酵素(KS)ドメイン
二次代謝産物の生合成に関与するポリケタイド合成酵素(PKS)に必要とされるドメインの一つ。通常は、二次代謝産物の生合成において炭素鎖伸長に関与する。KSはKetoshynthaseの略。
4.植物病原糸状菌
植物に対して病気を引き起こす糸状菌(かび)。
5.イネいもち病菌
学名はPyricularia oryzae(Magnaporthe oryzae)。病原性の糸状菌(かび)の一種。イネにいもち病を引き起こす。いもち病はイネの最重要病害であり、大幅な減収と食味の低下を引き起こす。
6.X線結晶構造解析
散乱されたX線を観測することで、物質の中の電子の分布、すなわち、物質の3次元構造を知る手法。
7.二次代謝産物
生物体を構成、維持する上で重要な物質を一次代謝産物と呼ぶ。一方、生育そのものには必要とされない代謝産物を二次代謝産物と呼び、抗生物質などが含まれる。
8.Alternaria(アルタナリア)
世界中に広く分布する植物病原糸状菌で、さまざまな穀物や果物などに被害を及ぼす。テヌアゾン酸以外にもさまざまなかび毒を生産する。
9.ポリケチド合成酵素(PKS)
ポリケチドを合成する多ドメイン酵素または酵素複合体。真正細菌や真菌、植物、少数の動物が持つ。ポリケチドは微生物や植物の二次代謝産物の中の主要なグループであり、さまざまな生理活性物質を含む。PKSはPolyketide synthaseの略。
10.非リボソーマルペプチド合成酵素(NRPS)
リボソームを用いず、ペプチドを生合成する酵素。非リボソームペプチドは細菌や真菌など微生物の二次代謝産物の中で、リボソームを経由せずに合成されるペプチドを指す。NRPSはnon-ribosomal peptide synthetaseの略。
11.ゲルろ過
タンパク質などを分子量の違いにより分離する方法。分子量に関する情報が得られる。
12.I型PKS
ポリケチド合成酵素(PKS)の一種であり、複数のドメインで構成される特徴を持つ。
13.α-ヘリックス
タンパク質の二次構造の一種で、バネのような右巻きらせんの形をしている。
14.オキサゾロマイシン
放線菌の一種が生産する二次代謝産物の一種。
15.ドッキングシミュレーション
主に小分子をあるタンパク構造に結合させ、結合力を推定するために使用するシミュレーション技術。
共同研究チーム
理化学研究所 環境資源科学研究センター
ケミカルバイオロジー研究グループ
研究員 尹 忠銖(ユン・チュンス)
専任研究員 本山 高幸(もとやま たかゆき)
研究嘱託 清水 猛(しみず たけし)
グループディレクター 長田 裕之(おさだ ひろゆき)
生命分子解析ユニット
ユニットリーダー 堂前 直(どうまえ なおし)
鳥取大学 工学部化学 バイオ系学科
大学院生(研究当時) 西本 一希(にしも とかずき)
准教授 日野 智也(ひの ともや)
教授 永野 真吾(ながの しんご)
研究支援
本研究は、農林水産省「農林水産業・食品産業科学技術研究推進事業」及び生物系特定産業技術研究支援センター「イノベーション創出強化研究推進事業」の研究課題「植物保護を目指した天然物ケミカルバイオロジー研究(研究統括:長田裕之)」などによる支援を受けて行われました。
原論文情報
Choong-Soo Yun, Kazuki Nishimoto, Takayuki Motoyama, Takeshi Shimizu, Tomoya Hino, Naoshi Dohmae, Shingo Nagano and Hiroyuki Osada, “Unique features of the ketosynthase domain in a nonribosomal peptide synthetase-polyketide synthase hybrid enzyme, tenuazonic acid synthetase 1”, Journal of Biological Chemistry, 10.1074/jbc.RA120.013105
発表者
理化学研究所
環境資源科学研究センター ケミカルバイオロジー研究グループ
研究員 尹 忠銖(ユン・チュンス)
専任研究員 本山 高幸(もとやま たかゆき)
グループディレクター 長田 裕之(おさだ ひろゆき)
鳥取大学 工学部 化学バイオ系学科
教授 永野 真吾(ながの しんご)
報道担当
理化学研究所 広報室 報道担当

