
2021-02-19 基礎生物学研究所
卵管は卵巣から放出された卵を取り込み、子宮へと輸送する生殖に必須の器官です。卵管の内側の細胞は多数の繊毛を持っており、この繊毛の向きが揃い、同じ方向に運動することが卵の輸送に重要です。卵管内の繊毛の向きが揃わなくなると、卵を輸送する能力が低下し、不妊の要因になることが知られています。基礎生物学研究所の宇佐美(松川) 文子大学院生と藤森 俊彦教授らは、理化学研究所の竹市 雅俊博士、ベルギー ルーヴァン・カトリック大学のFadel Tissir博士との共同研究により、卵管の内側にある多繊毛細胞の繊毛の向きが、CAMSAP3とCELSR1という2つのタンパク質を介して細胞内、細胞間で揃えられることを明らかにしました。本成果はJournal of Cell Science誌に掲載されました。

図1:繊毛の向きが細胞内、細胞間で揃えられる仕組みのまとめ
繊毛基部に局在するCAMSAP3によって細胞内において繊毛の向きが揃えられる。また、細胞境界部に局在するCELSR1によって細胞間で繊毛の向きが揃えられる。
【研究の背景】
卵管は卵巣から放出された卵を取り込み、子宮へと輸送する生殖に必須の器官です。また、卵管内では卵が精子と出合い受精すること、さらに受精卵の細胞分裂など子宮に至るまでの間に発生が進むことが知られています。卵管の内側の上皮は分泌細胞と1細胞当たり平均200本の繊毛を持つ多繊毛細胞からなり、卵巣に近い卵管の漏斗と呼ばれる領域では多繊毛細胞が上皮の約80%を占めています。繊毛が細胞内、細胞間で同じ方向に運動することにより、卵巣から子宮へと卵は輸送されます(図2)。卵管内の繊毛の向きが揃わなくなると、卵を輸送する能力が低下し、不妊の要因になります。卵管のほかに、気管や脳室にも多繊毛細胞があり、繊毛の方向性を持った運動により異物の除去や物質の輸送が可能となります。このような器官の多繊毛の向きがどのようにして細胞内、細胞間で揃えられるかは、十分に理解が進んでいませんでした。
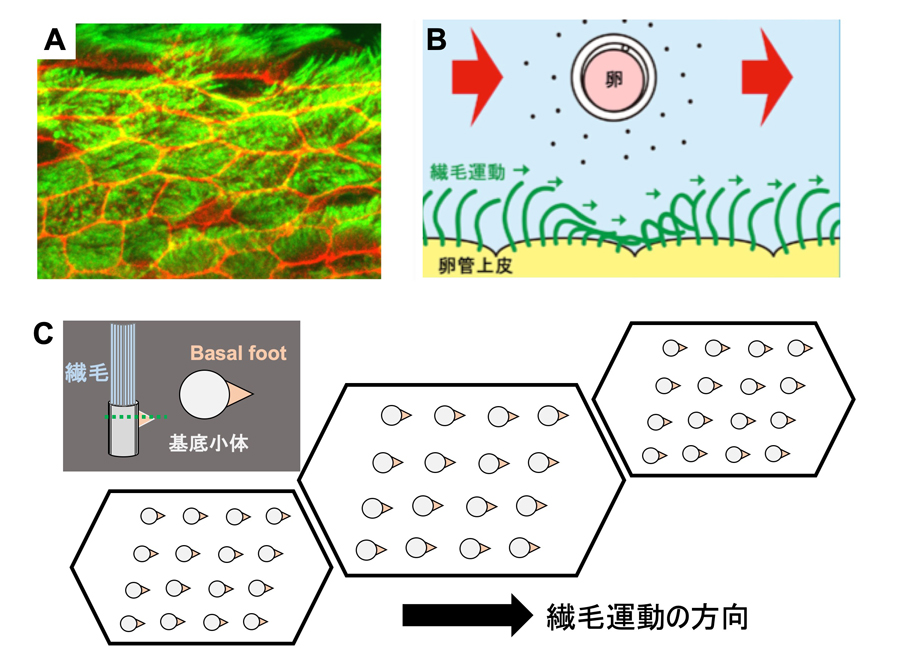 図2:卵管の内側にある繊毛によって卵が一方向に輸送される
図2:卵管の内側にある繊毛によって卵が一方向に輸送される
A:卵管上皮の繊毛(緑)と細胞形態(赤)を可視化した像(上皮を真上から観察)。1つの多繊毛細胞は約200本の繊毛を有する。
B:卵が輸送される模式図。卵管上皮の多繊毛が同じ方向に揃って運動することによって、卵は卵管内を輸送される。
C:多繊毛細胞における繊毛の模式図。繊毛の根元には基底小体と呼ばれる構造があり、基底小体から三角状に突出した構造(basal foot)が見られ、basal footの方向と繊毛の運動方向は一致する。今回、基底小体とbasal footを染色することにより、卵管組織内の繊毛の向きを調べた(手法の詳細は図3参照)。
【研究の成果】
藤森研究室の先行研究でCELSR1と呼ばれるタンパク質を失ったマウスでは、卵管の繊毛運動の方向が乱れ、卵の輸送能力が失われることを明らかにしていました。しかし、CELSR1 がどのようにして繊毛の方向を制御するかはわかっていませんでした。そこで、Celsr1を欠損したマウス(Celsr1変異体)の多繊毛細胞を正常なマウス(野生型マウス)と比較して詳細に調べることにより、卵管の繊毛の向きが細胞内、細胞間で揃う仕組みを明らかにすることをめざしました。
研究グループは、超解像顕微鏡観察により、卵管上皮の広い範囲でそれぞれの繊毛の向きを判定できる手法を開発しました(図3)。
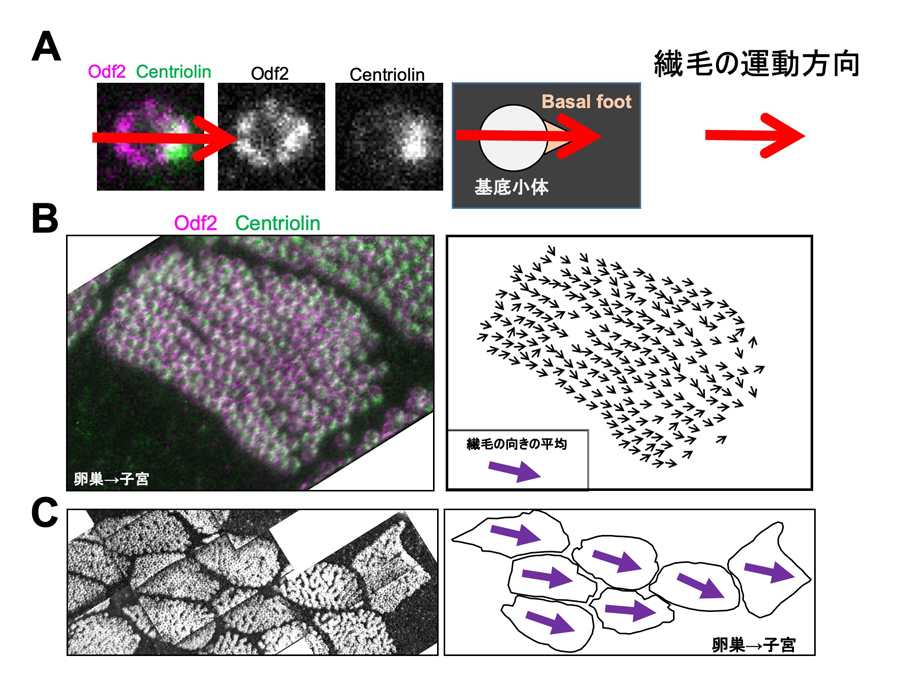 図3:超解像顕微鏡観察により、繊毛の向きを判定する手法を開発
図3:超解像顕微鏡観察により、繊毛の向きを判定する手法を開発
研究グループは今回、超解像顕微鏡を用いて細胞内および細胞間の繊毛の向きを判定する手法を開発した。
A:繊毛の基底小体とbasal footを染色し、繊毛の向きを判定した。リング状の基底小体(Odf2染色、マゼンタ)に対して偏って存在するbasal foot(Centriolin染色、緑)が楕円状のシグナルとして観察される。リングの中心と楕円の中心を結ぶ方向を繊毛の運動方向として定義する(赤矢印)。
B:野生型マウスの1つの細胞の染色像(左の図)、繊毛の向き判定(右の図)の例であり、それぞれの黒い矢印が繊毛1つずつの向きを示す。細胞内で繊毛の向きが揃って観察される。1つの細胞内の繊毛の向き(黒い矢印)から平均の角度(紫色の矢印)を計算した。
C:隣接する複数の細胞間で繊毛の向きの平均を比べるため、複数の写真を貼り合わせた。野生型マウスの観察例では、隣接する細胞間で、矢印の方向が揃っている。
開発した手法で多繊毛細胞の繊毛の向きを調べたところ、野生型マウスではそれぞれの細胞内においても細胞間においても繊毛は同じ方向を向いているのに対し、Celsr1変異体では細胞内では繊毛の向きはあまり乱れておらず、一方で細胞間では大きく乱れていることが明らかになりました。次に、卵管の多繊毛細胞で微小管の分布を調べました。野生型マウスでは、それぞれの細胞の子宮側細胞境界に濃縮している微小管、細胞の表面(アピカル面)付近で繊毛の基部の間に縞状に分布する微小管、そして繊毛基部から細胞の基底部に向かう微小管が見られました。Celsr1変異体では微小管の細胞境界への濃縮に異常が見られました。約75%の細胞では微小管の濃縮が見られましたが、濃縮している方向は細胞によって異なっていました。残りの約25%の細胞では微小管の濃縮が不明瞭でした。微小管の濃縮が見られたCELSR1欠損細胞では、微小管が濃縮している方向に繊毛の向きが揃っている傾向がありました(図4)。これらの結果から、CELSR1を欠損した細胞においても、個々の細胞内で繊毛の向きを揃える機構が維持されていることが示唆されました。野生型マウスにおいては、CELSR1が微小管の濃縮方向を細胞間で子宮側に揃えることによって、繊毛の向きが揃えられていると考えられます。
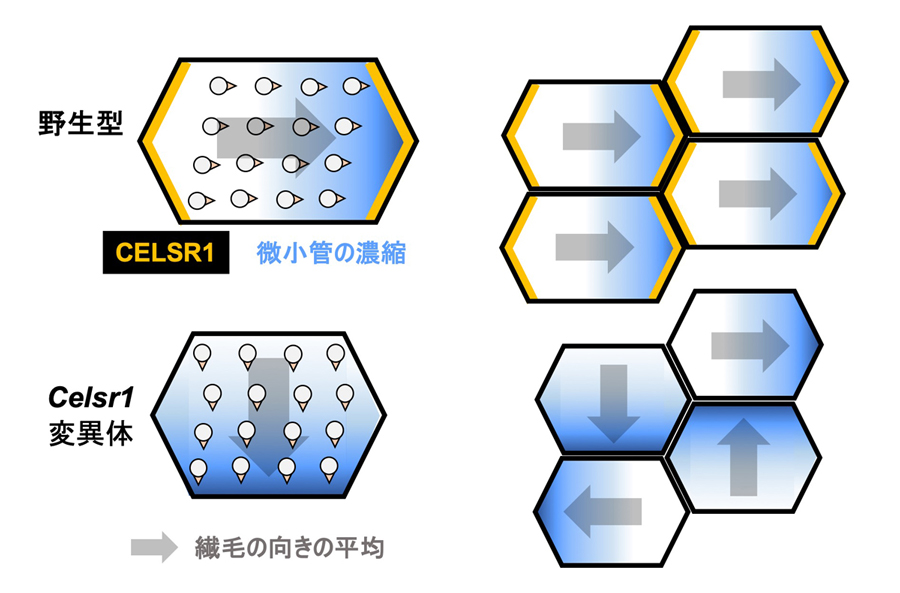 図4:CELSR1(膜貫通型の平面内細胞極性因子)を失った変異体でみられた異常のまとめ
図4:CELSR1(膜貫通型の平面内細胞極性因子)を失った変異体でみられた異常のまとめ
CELSR1は、それぞれの細胞の卵管側と子宮側の細胞辺に存在する(黄色)。CELSR1を失った細胞では、個々の細胞内での繊毛は大きく乱れることはない(下段左)。一方、隣接する細胞間で繊毛の向きは揃っていない(下段右)。また、細胞境界近傍に微小管の濃縮が見られ(青)、その方向は野生型マウスでもCelsr1変異体でも繊毛の向きと一致する。
次に、微小管が繊毛の向きの制御に関わる可能性を検証するため、微小管のマイナス端に結合するCAMSAP3というタンパク質に注目しました。野生型マウスにおいてCAMSAP3は、それぞれの繊毛の根元の領域で子宮側に偏って存在していることを見いだしました(図5)。
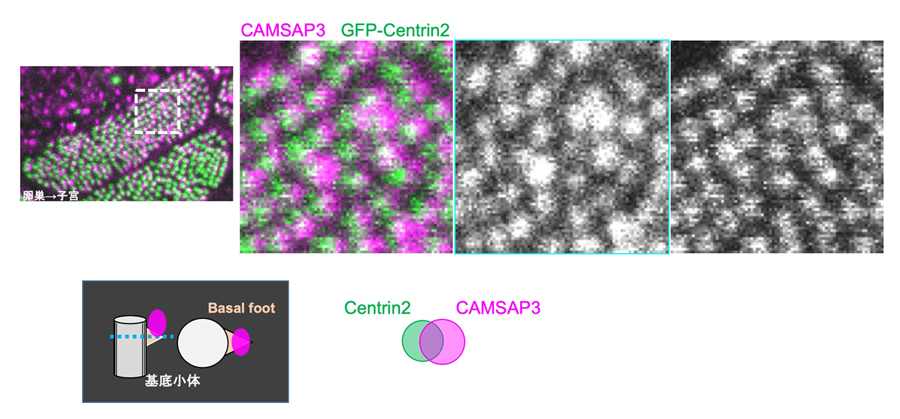 図5:CAMSAP3は繊毛基部に偏って存在する
図5:CAMSAP3は繊毛基部に偏って存在する
上段:免疫染色によりCAMSAP3(マゼンタ)を観察した。繊毛基部の基底小体(マーカーとしてGFP-Centrin2(緑)を用いた)付近でCAMSAP3は子宮側に偏って存在していることがわかった。
下段:CAMSAP3の局在の模式図。繊毛基部で、basal footと同様に子宮側に存在する。
そこで、繊毛の向きの制御にCAMSAP3が関与するか調べる為に、CAMSAP3に変異を持つマウス(Camsap3変異体)の卵管多繊毛細胞を調べました。このマウスの卵管では、細胞内で繊毛の向きが乱れ、細胞によっては向きがランダムになっていました(図6)。このことから、CAMSAP3が細胞内で繊毛の向きを揃えるのに必要であることがわかりました。さらに、CAMSAP3に変異を持つマウスでは、細胞のアピカル面付近に存在する微小管の分布に異常がみられました(図7)。野生型マウスにおいては、CAMSAP3が細胞のアピカル面で繊毛基部付近の微小管を制御することによって、繊毛が微小管の濃縮する方向を向くように揃えられていると考えられます(図8)。しかし、Camsap3変異体でも野生型マウスと同じようにCELSR1が細胞境界に存在し、微小管も子宮側の細胞境界に濃縮していたことから、細胞全体の方向を細胞間で揃える機構には異常がないことが示唆されました。また、CELSR1を欠損した多繊毛細胞においても、CAMSAP3は野生型マウスと同じ局在を示したことから、CELSR1はCAMSAP3の制御には直接関わらないことが示唆されました。
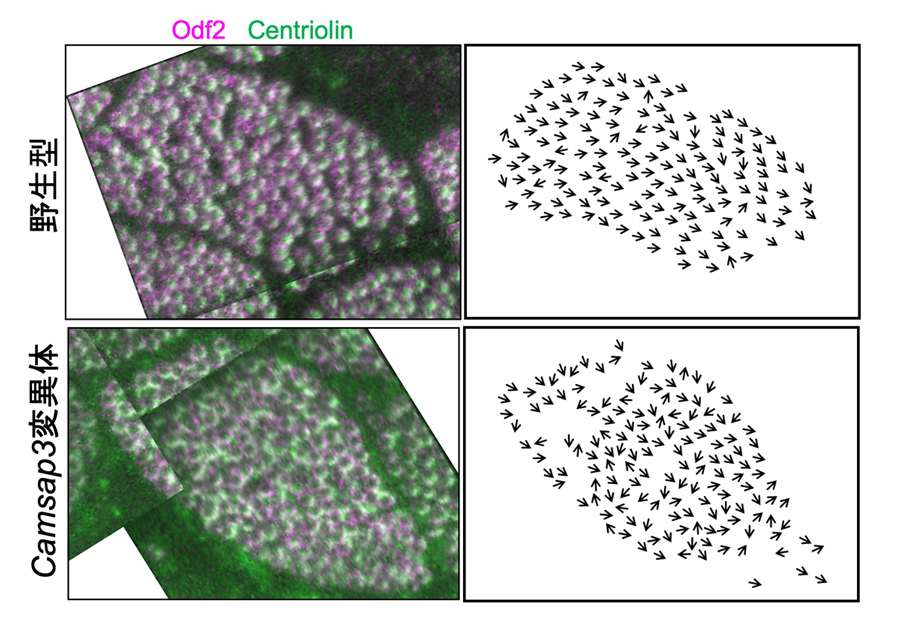 図6:Camsap3変異体においては、細胞内で繊毛の向きが乱れる
図6:Camsap3変異体においては、細胞内で繊毛の向きが乱れる
超解像顕微鏡を用いて繊毛の向きを判定すると、Camsap3 変異体(下段)ではそれぞれの繊毛の向き(矢印で示す)が1つの細胞の中で揃っていない。(左の図は、シグナルが重なるように撮影した複数の写真を貼り合わせている)
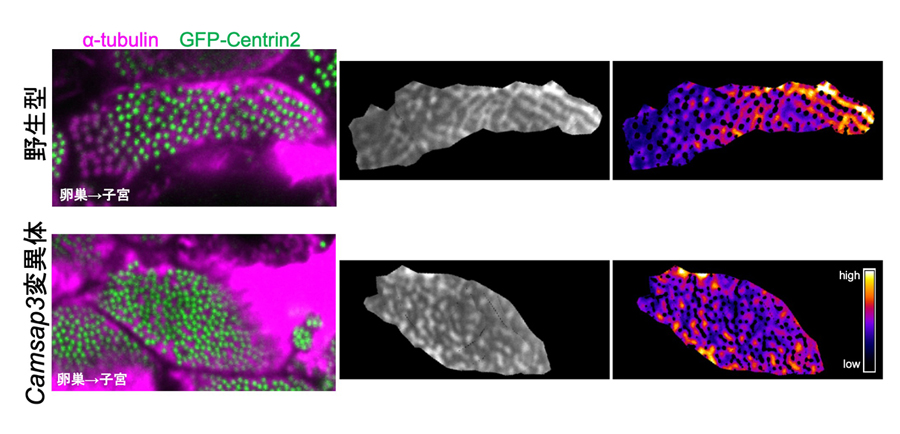 図7:Camsap3変異体では繊毛基部付近の微小管の分布が乱れる
図7:Camsap3変異体では繊毛基部付近の微小管の分布が乱れる
野生型(上段)においては、繊毛基部(左パネルの緑)付近で微小管(左パネルのマゼンタ; 中央・右パネル)がストライプ状に見られ、子宮側(右側)で濃いシグナルが見られる。一方、Camsap3変異体においては、ストライプ構造は形成されず、細胞内でシグナルの強弱も曖昧である。右のパネルでは中央のパネル内のシグナルの強弱を色分けして示している。
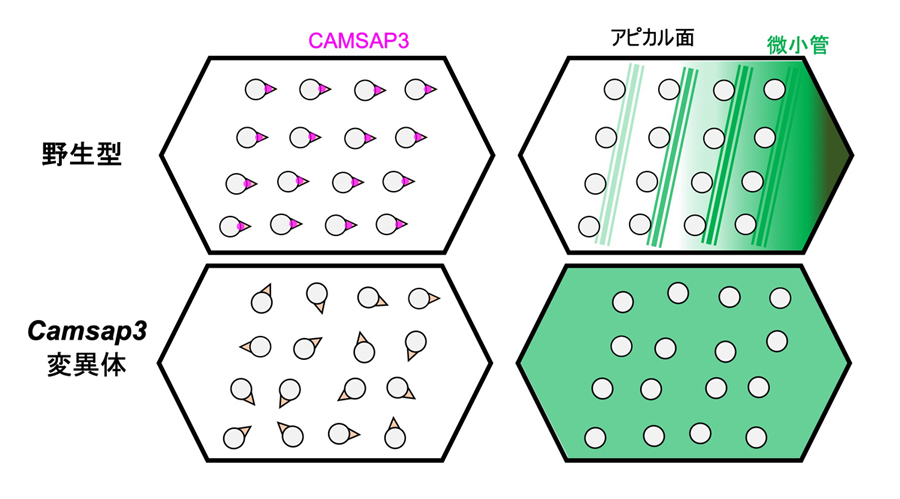 図8:Camsap3変異体における異常のまとめ
図8:Camsap3変異体における異常のまとめ
Camsap3変異体では、細胞内で繊毛の向きが揃わず(下段、左)、繊毛基部を取り巻く微小管の配置も異常となる。
以上の結果から、CAMSAP3とCELSR1によって、繊毛の向きが細胞内、細胞間で揃えられる仕組みが明らかとなりました(図9)。これらの2つの仕組みが連携して、組織内全体で繊毛の向きが揃えられていると考えられます。
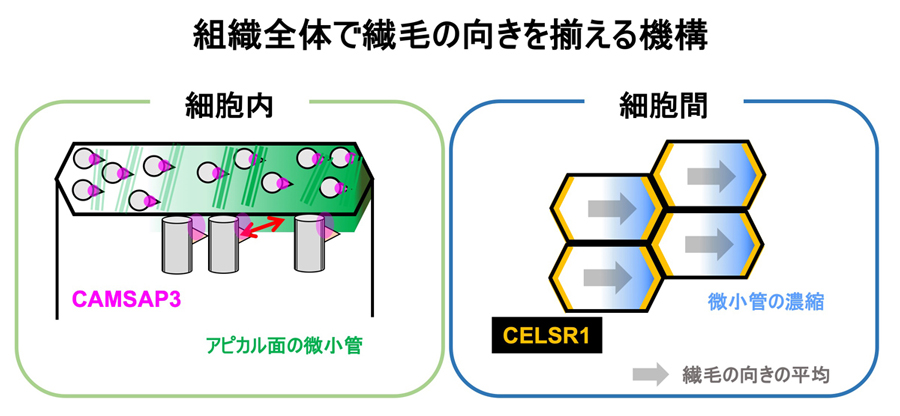 図9:繊毛の向きが細胞内、細胞間で揃えられる仕組みのまとめ(図1の再掲)
図9:繊毛の向きが細胞内、細胞間で揃えられる仕組みのまとめ(図1の再掲)
繊毛基部に局在するCAMSAP3によって細胞内において繊毛の向きが揃えられる。また、細胞境界部に局在するCELSR1によって細胞間で繊毛の向きが揃えられる。
【今後の展望】
今回の研究により、CAMSAP3とCELSR1を介して細胞内、細胞間で繊毛の向きを揃える仕組みが明らかになりましたが、この2つの仕組みが微小管を介してどのように相互作用しているかはまだわかっていません。宇佐美大学院生は、「繊毛の向きの制御だけでなく、卵管の長軸方向に沿った細胞の伸長や上皮のヒダ構造など、多階層にわたる卵管細胞の極性がどのようにして制御されているのかを明らかにしたい。更に、卵管以外の器官においても膨大な数の細胞がどのようにして秩序のある構造を作り出しているのか研究を進めていきたい」と話しています。
【発表雑誌】
雑誌名: Journal of Cell Science
掲載日: 2021年1月19日
論文タイトル: Intercellular and intracellular cilia orientation is coordinated by CELSR1 and CAMSAP3 in oviduct multiciliated cells
著者:Fumiko Matsukawa Usami, Masaki Arata, Dongbo Shi, Sanae Oka, Yoko Higuchi, Fadel Tissir, Masatoshi Takeichi, and Toshihiko Fujimori
DOI: https://doi.org/10.1242/jcs.257006
【研究グループ】
本研究は、基礎生物学研究所 初期発生研究部門の宇佐美(松川) 文子(総合研究大学院大学 生命科学研究科 基礎生物学専攻 大学院生)、新田昌輝研究員(名古屋大学 特任助教)、藤森俊彦教授らが中心となって、ドイツ ハイデルベルグ大学 石東博博士(元 初期発生研究部門 NIBBリサーチフェロー)、ベルギー ルーヴァン・カトリック大学のFadel Tissir博士、理化学研究所の竹市 雅俊博士との共同研究として行われました。
【研究サポート】
本研究は、文部科学省 日本学術振興会の科研費(25291054, 15H01220, 17H03689, 16H06280, 19K16153)、国立研究開発法人 科学技術振興機構 戦略的創造研究推進事業(JPMJCR1654)のサポート、および基礎生物学研究所からのサポートを受けて実施されました。宇佐美(松川)文子大学院生は、総合研究大学院大学の国際インターンシップ事業のサポートにより、Fadel Tissir博士の研究室を訪問しました。
【本研究に関するお問い合わせ先】
基礎生物学研究所 初期発生研究部門
教授: 藤森 俊彦 (フジモリ トシヒコ)
【報道担当】
基礎生物学研究所 広報室

