2023-04-25 東京大学
佐野 文哉(生物科学専攻 博士課程)
赤坂 浩明(生物科学専攻 修士課程)
志甫谷 渉(生物科学専攻 助教)
濡木 理(生物科学専攻 教授)
発表のポイント
- Gタンパク質共役受容体の効率的な構造解析を可能にする発現・精製系であるFusion-Gシステムを確立し、エンドセリン1の結合により活性化した状態の受容体の立体構造を高分解能で決定しました。
- 得られた立体構造を不活性状態の構造と比較することにより、エンドセリン受容体がもつユニークなGタンパク質活性化機構を明らかにしました。
- 本研究はエンドセリン受容体を標的とした薬剤の合理的な開発に向けた構造基盤を提供します。

本研究の概要図
発表概要
今回、東京大学大学院理学系研究科の佐野文哉大学院生、赤坂浩明大学院生、志甫谷渉助教、濡木理教授らのグループは、難易度の高い構造解析に向けて新規に確立したFusion-Gシステムと呼ばれる手法を用いて、クライオ電子顕微鏡(注1)によって、エンドセリン1(注2)、エンドセリンB型受容体、およびGiタンパク質三量体(注3)からなるシグナル伝達複合体の立体構造を決定しました。
エンドセリン1(ET-1)は血流調節作用や神経保護作用を持つ生理活性ペプチドとして知られています。ET-1を受容するエンドセリンB型受容体(ETB)は血管内皮細胞や神経細胞で発現しており、Gタンパク質共役受容体(GPCR)(注4)と呼ばれる膜タンパク質の一群に属します。細胞外側でET-1を受容したETBは構造変化を引き起こし、細胞内側で異なる複数のGタンパク質を介してシグナルを伝達します。このようにしてET-1とETBによるシグナル伝達系は、生体内における局所的な血流の調節や反応性アストロサイトの形成を制御しており、抗がん剤の効率的な送達法や優れた神経保護薬の開発に向けて注目を集めています。しかし、ET-1の結合によるETBの活性化機構は未解明であり、これらを標的とした合理的な薬剤設計はこれまで困難でした。
本研究で得られた立体構造から、ET-1の結合によって引き起こされるETB独自の活性化機構の詳細が明らかになりました。さらに他のGPCRの活性化機構と比較することで、異なる複数のGタンパク質と結合可能であるというETBの特性を説明することに成功しました。
本研究は合理的な薬剤の設計に向けた構造情報を提供し、がんや神経変性疾患に対する創薬研究に貢献すると期待されます。
発表内容
〈研究の背景〉
Gタンパク質共役受容体(GPCR)は細胞膜上に存在し、リガンドと結合すると活性型構造へと変化します。そして細胞内に存在するGタンパク質三量体と結合し、活性化させることで細胞内へとシグナルを伝達します。ETBは多数存在するGPCRのうちの1つであり、生理活性ペプチドであるET-1を受容することで生体内における局所的な血流の調節や反応性アストロサイトの形成を制御していることが知られています。このような生理作用を持つことから、ET-1とETBによるシグナル伝達系は抗がん剤の効率的な送達法や優れた神経保護薬の開発に向けて研究が進められてきました。しかし、これまで報告されたETBの立体構造は受容体単体の不活性状態および部分的な活性状態のみに留まっていました。ETBでは、ほぼ全てのGPCRで保存されている、Gタンパク質の活性化に重要なモチーフが保存されていないほか、全ての種類のGタンパク質を活性化できるといった特徴を持つことが知られています。このような独自の特徴を持つETBのGタンパク質活性化機構の詳細は全く分かっていませんでした。
〈研究の内容〉
本研究ではまず、構造解析に向けた効率的なGPCR-Gタンパク質複合体の発現・精製系であるFusion-Gシステムを確立しました。Fusion-Gシステムは受容体のC末端にNanoBiTとよばれる酵素の大きな断片(LgBiT)を融合し、残った小さな断片(HiBiT)をGβサブユニットのC末端に融合させ、さらにリンカーを介してGαサブユニットとGγサブユニットを融合させる手法です。これにより、安定性の低いGPCR-Gタンパク質複合体を発現・精製することが可能となりました。本研究ではFusion-Gシステムを適用することにより、ET-1によって活性化されたETBとGiタンパク質三量体との複合体の立体構造をクライオ電子顕微鏡法によって決定しました(図1)。
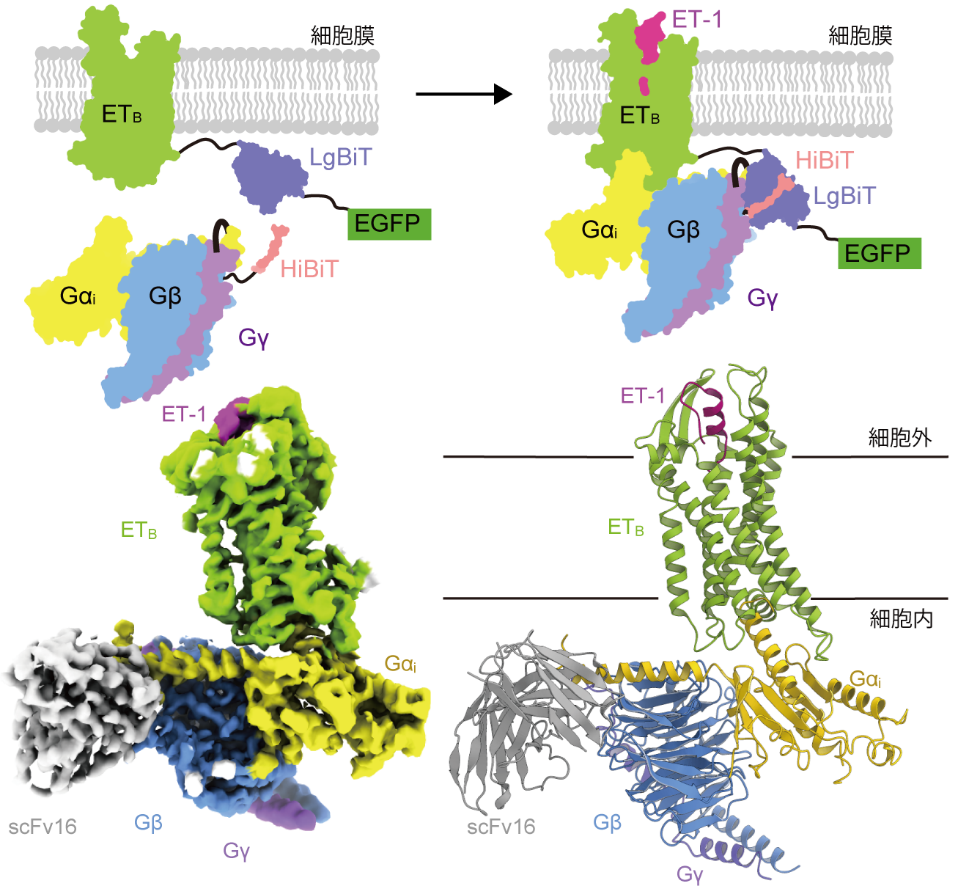
図1:Fusion-Gシステムの概要 (上)、およびETB-Gi複合体の構造 (下)
Fusion-Gシステムは2つの技術を組み合わせたGPCRの発現・精製系です。1つがNanoBiTとよばれる酵素の大きな断片(LgBiT)を受容体のC末端に融合し、残った小さな断片(HiBiT)をGβのC末端に融合させる手法、もう1つがリンカーを介してGαサブユニットとGγサブユニットを融合させる手法です。これらの戦略を組み合わせることで、安定性の低いGPCR-Gタンパク質複合体を容易に調製することが可能となりました(上)。クライオ電子顕微鏡によって得られた密度マップと原子モデル。ET-1の結合により活性化したETBがGiタンパク質三量体と複合体を形成し、構造認識抗体scFv16によって安定化されていることが分かります (下)。
不活性状態および部分的な活性状態の構造と比較することにより、ET-1の結合によるETBの活性化機構の詳細が明らかになりました。細胞外側においてETBは他のGPCRと共通の活性化機構を持つことが分かりました。ET-1の結合により、ETBのリガンド結合ポケット底部にあるトリプトファン残基が下方に押し込まれます。それにより、近傍に存在する疎水性の残基群が細胞内方向へと回転するような構造変化を引き起こします。最終的に細胞内側で膜貫通ヘリックス6番(TM6) が大きく外側へと開き、Gタンパク質と結合するためのポケットが形成されます。一方で細胞内側に着目すると、ETBの活性化機構は他のGPCRとは大きく異なっていました。多くのGPCRでは活性化に際してTM7が内側にスライドすることが知られています。これはTM5とTM7に存在する広く保存されたチロシン残基が、活性化に伴い水を介した水素結合を形成するためです。しかしながらETBではTM7のチロシンは保存されていないため水素結合が形成されません。結果としてETBではTM7の内側方向へのスライドが起こらず、代わりに下方向へのスライドが観察されました(図2)。
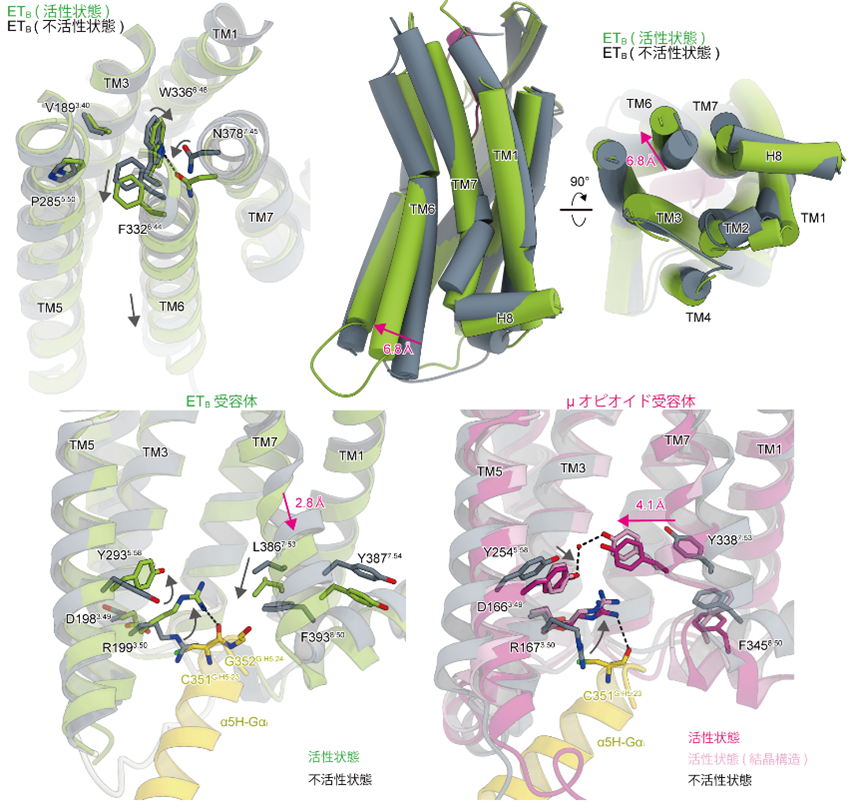
図2:既に報告されている不活性状態と重ね合わせた図。
細胞外側ではGPCRの活性化における典型的な構造変化が観察されました(左上)。ET-1の結合により疎水性の残基群が細胞内側へと押し下げられ、最終的に細胞内側で膜貫通ヘリックス6番(TM6)が外側へ開くような構造変化を起こします(右上)。一方で、細胞内側では膜貫通ヘリックス7番(TM7)が下方向へシフトする特徴的な構造変化が観察されました(左下)。代表的な他のGPCRでは、活性化に伴うTM7のシフトは内側方向であることが分かります(右下)。
このようなユニークな活性化機構をもつため、ETBは他のGPCRとは異なる形式でGタンパク質と結合します。多くのGPCRでは受容体コアとGタンパク質C末端の接触部分が最も主要な相互作用界面であることが知られています。ETBの細胞内側ポケットに対するGタンパク質C末端の相対位置は主要なGPCR-Gi複合体の中で最も浅く、それほど強い相互作用を形成していないと考えられます。この部位での相互作用の弱さを補完するように、ETBの細胞内ループ2番やC末端がGiと相互作用していることが分かりました。このようにETBではGタンパク質C末端のアミノ酸認識がそれほど厳密ではないことが分かります。こうした独自のGタンパク質認識機構は、ETBのもつ「異なる複数のGタンパク質と結合可能である」という特性を説明していると考えられます(図3)。
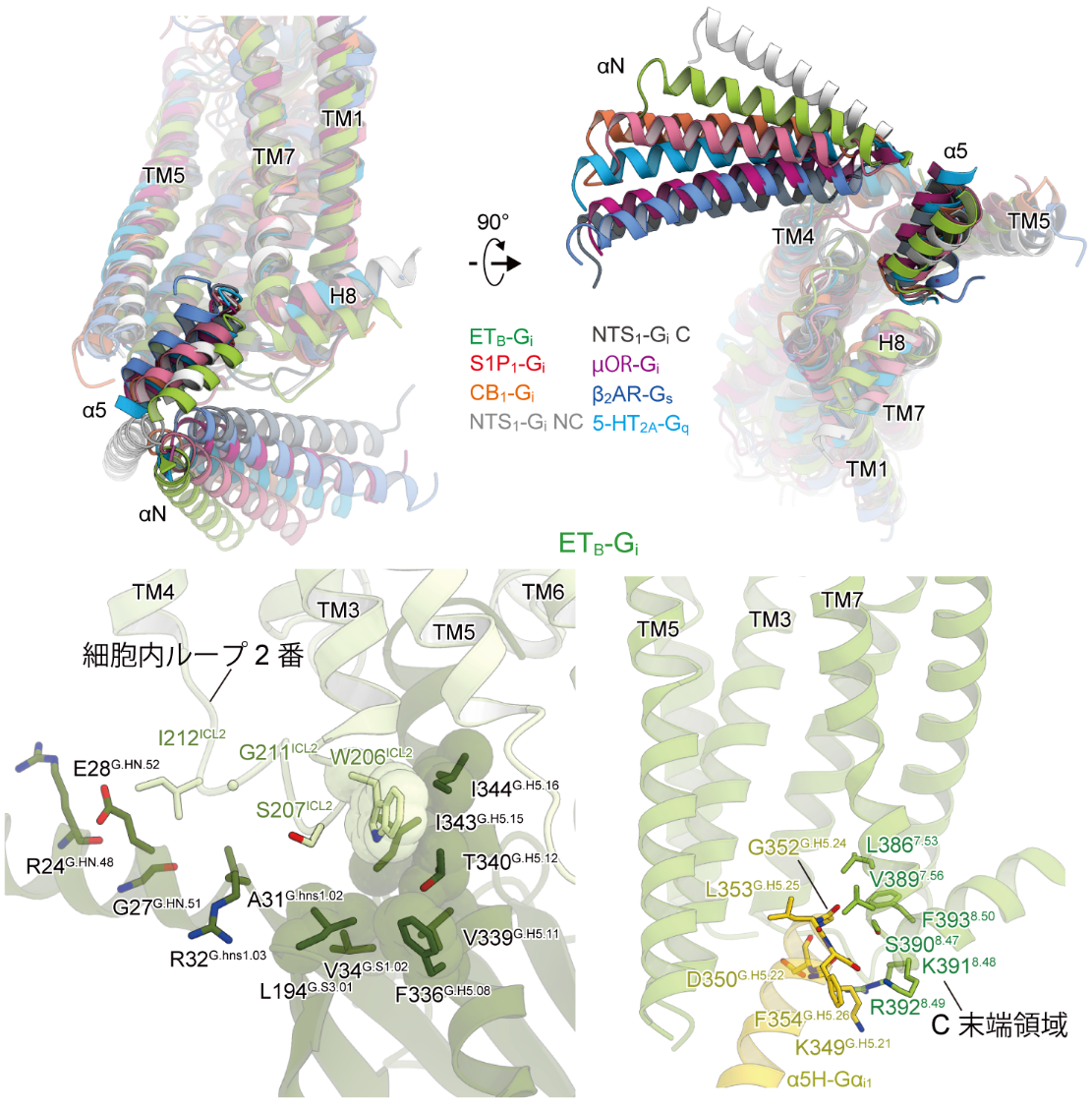
図3:Gi結合様式の比較およびETBの特徴的な相互作用
細胞外側ではGPCRの活性化における典型的な構造変化が観察されました(左上)。ET-1の結合により疎水性の残基群が細胞内側へと押し下げられ、最終的に細胞内側で膜貫通ヘリックス6番(TM6)が外側へ開くような構造変化を起こします(右上)。一方で、細胞内側では膜貫通ヘリックス7番(TM7)が下方向へシフトする特徴的な構造変化が観察されました(左下)。代表的な他のGPCRでは、活性化に伴うTM7のシフトは内側方向であることが分かります(右下)。
〈今後の展望〉
本研究で確立したFusion-Gシステムは他のGPCR複合体の構造解析にも応用できる可能性があり、構造生物学研究のさらなる発展に寄与すると考えられます。また本研究はETBを標的とした合理的な薬剤の設計に向けた構造情報を提供し、がんや神経変性疾患に対する創薬研究に貢献すると期待されます。
〈関連のプレスリリース〉
「がんや肺炎にかかわるLPA受容体の活性型構造の解明」(2022/9/20)
https://www.s.u-tokyo.ac.jp/ja/press/2022/8046/
「最後に残されたβアドレナリン受容体の立体構造を解明」(2021/7/26)
https://www.s.u-tokyo.ac.jp/ja/press/2021/7484/
論文情報
- 雑誌名
eLife論文タイトル
Cryo-EM structure of the endothelin-1-ETB-Gi complex著者
Fumiya K. Sano, Hiroaki Akasaka, Wataru Shihoya*, Osamu Nureki* *共同責任著者DOI番号
10.7554/eLife.85821
研究助成
本研究は、日本学術振興会における科学研究費助成事業の基盤S「生体環境でのGPCRの構造ダイナミクス」(課題番号:21H05037 研究開発代表者:濡木 理)や基盤B「クライオ電子顕微鏡法を用いたGPCR創薬研究」(課題番号:22H02751 研究開発代表者:志甫谷 渉)などの一環で行われました。また、本研究は、AMED「創薬等ライフサイエンス研究支援基盤事業」および「革新的先端研究開発支援事業インキュベートタイプ」の一環として、放射光施設などの大型施設の外部開放を行うことで優れたライフサイエンス研究の成果を医薬品等の実用化につなげることを目的とした「創薬等先端技術支援基盤プラットフォーム(BINDS)」の支援により行われました。
用語解説
注1 クライオ電子顕微鏡
液体窒素(-196℃)冷却下でタンパク質などの分子に対して電子線を照射し、試料の観察を行うための装置です。タンパク質や核酸をはじめとする生体高分子の像を多数撮影することで立体構造の決定を行う、単粒子解析法に用いられます。↑
注2 エンドセリン1(ET-1)
ET-1は、主に内皮で産生される21アミノ酸の生理活性ペプチドであり、血管の恒常性維持に重要な役割を担っています。ET-1は、Gタンパク質共役型受容体(GPCR)のクラスAに属するETAおよびETBという2つの受容体サブタイプを通じてシグナルを伝達します。これらのエンドセリン受容体は、成長、生存、浸潤、血管新生に必要となる細胞プロセスの制御を行います。また、ETB受容体を介したGiシグナルは細胞骨格の再編成と細胞接着依存性の増殖をもたらし、反応性アストロサイトの誘導と神経保護を促進することが知られています。↑
注3 Gタンパク質、Giタンパク質三量体
Gタンパク質は、細胞内情報伝達に関わるGTP結合タンパク質であり、Gα、Gβ、Gγサブユニットの三量体により構成されます。活性化されたGタンパク質共役型受容体(GPCR)と結合したGタンパク質三量体では、GDP-GTP交換反応が起き、GαとGβ-Gγの二つに解離します。解離したサブユニットは下流のシグナル伝達因子と結合し活性化させることで、細胞にさまざまなシグナル応答を生じさせます。Gαサブユニットは大きくGs、Gi、Gq/11、G12/13の4種類に分別され、特にGiタンパク質三量体は、下流でアデニル酸シクラーゼの活性を阻害することで、抑制性のシグナルを伝達します。↑
注4 Gタンパク質共役受容体(GPCR)
GPCRは細胞膜に発現する膜タンパク質で、膜を貫通する7本のαヘリックスで構成されています。膜受容体タンパク質のなかで最大のファミリーを形成し、細胞外領域で特定のリガンドが結合することで活性化し、細胞内のGタンパク質三量体を活性化することでシグナルを伝達します。GPCRは生体内における多様なプロセスの調節にはたらいており、既承認薬の30%以上がGPCRを標的としています。
が法律的に「人」とみなされうることを指摘")