2019-07-11 理化学研究所,京都大学
理化学研究所(理研)生命機能科学研究センター非対称細胞分裂研究チームの河野夏鈴大学院生リサーチ・アソシエイト(京都大学大学院生命科学研究科博士課程3年)、松崎文雄チームリーダー(京都大学大学院生命科学研究科教授)らの研究グループ※は、細胞極性[1]の形成に働くタンパク質複合体によって細胞に非対称性が生じる際の基本的なプロセスを解明しました。
本研究成果は、細胞極性が関わる器官形成・再生や、細胞極性の乱れによって生じるがん化メカニズムの詳細な理解に貢献すると期待できます。
「細胞極性」は、細胞に方向性を与える細胞の基本的な性質で、器官や組織を正常に形成するために欠かせない要素です。細胞に極性を作り出すシステムの中でも、Parタンパク質[2]の複合体(Par複合体)による極性形成は、線虫からヒトまで保存され、さまざまな組織で機能する最も一般的なシステムです。しかし、Par複合体の振る舞いや特徴などはモデル生物や組織ごとに異なるため、その普遍的なプロセスは不明でした。
今回、研究グループは、Parタンパク質の発現量を操作することで、通常は極性を持たないショウジョウバエ胚由来の培養細胞上に、人為的に非対称的なタンパク質の分布を作り出すことに成功しました。この細胞極性の再構成により、細胞が非対称性を生み出す典型的なプロセスを高解像度で観察可能となり、Par複合体が3段階の動的な凝集状態を経ることが分かりました。これらの凝集体は、ショウジョウバエの個体内の細胞でも存在することが確認されたことから、Par複合体が働く細胞極性の形成に一般的な過程と考えられます。また、凝集状態の最終段階である島状の凝集体を超解像顕微鏡[3]で観察した結果、この凝集体はセグメント状の構造単位が網目状に集まった構造を持つことが分かりました。
本研究は、英国のオンライン科学雑誌『eLife』(6月7日付け)に掲載されました。

図 Par複合体(緑)の凝集により培養細胞で非対称性が生まれるまでの概要図
※研究グループ
理化学研究所 生命機能科学研究センター
非対称細胞分裂研究チーム
大学院生リサーチ・アソシエイト 河野 夏鈴(こうの かりん)
(京都大学大学院 生命科学研究科 博士課程3年)
研究員(研究当時) 吉浦 茂樹(よしうら しげき)
特別研究員(研究当時) 藤田 生水(ふじた いくみ)
専門職研究員 下向 敦範(したむかい あつのり)
チームリーダー 松崎 文雄(まつざき ふみお)
(京都大学大学院 生命科学研究科教授)
フィジカルバイオロジー研究チーム
チームリーダー 柴田 達夫(しばた たつお)
細胞極性統御研究チーム
チームリーダー 岡田 康志(おかだ やすし)
背景
細胞の成分が非対称に分布し、方向性を持つことを「細胞極性」と呼びます。この細胞極性は、器官や組織の形成をはじめとした多様な現象に関わる細胞の基本的な性質です。細胞極性が乱れると、個体発生の異常や組織のがん化などが引き起こされるため、細胞極性は、発生期や成体を問わず個体にとって不可欠な細胞の特性です。
細胞に極性を作り出すさまざまなシステムの中でも、Parタンパク質の複合体(Par複合体)からなる極性形成システム(Par極性システム)は、線虫やショウジョウバエからヒトまで保存されているだけでなく、受精卵、神経幹細胞[4]や上皮細胞[5]などさまざまな組織で機能する最も一般的な極性形成システムです。Par極性システムの働きにより細胞が極性を作るとき、Par複合体の主要成分であるPar3-Par6-aPKC(非定型プロテインキナーゼC)が、細胞膜直下(細胞表層と呼ばれる)で非対称に分布します。このときのPar複合体の振る舞いや特徴などは、さまざまなモデル生物や組織で詳しく調べられてきました。
それにより明らかになったことは、Par複合体の振る舞いが、細胞極性と協調する他の生命現象に密接に関連しており、モデル生物や組織ごとに異なっていることです。このような生体におけるPar複合体の振る舞いの多様性は、Par極性システムの典型的な仕組みを明らかにすることを困難にしていました。
Par極性システムの普遍的な形成過程を観察するには、Par複合体と協調する生命現象の影響を取り除き、さらに観察が容易な実験条件を整備する必要があります。そこで研究グループは、通常は極性を持たない培養細胞を操作し、人為的にPar複合体の非対称性を培養細胞で作り出すことで、Par極性システムの極性形成で典型的な過程や原理を明らかにすることを目指しました。
研究手法と成果
Par複合体が極性を形成するときは、細胞表層で働きます。そこで研究グループは、Par複合体が細胞質内に均質に分布し、極性を持たないショウジョウバエ胚由来の培養細胞を用いて、Par複合体が細胞表層で非対称性な分布を形成する条件を探しました。その結果、Par複合体を構成するタンパク質の一つであるPar3の発現量を上昇させると、Par複合体の分布が、細胞質での一様な分布から細胞表層での非対称な分布へと変化することが分かりました(図1)。つまり、Par極性を持たない培養細胞に、Par極性を作り出すことに成功しました。また、培養細胞のPar複合体の分布の非対称性は、反応拡散系モデル[6]という生体内でよく見られるメカニズムに基づいて作り出されることを示唆する証拠を得ました。
次に、Par複合体の細胞質における均質な分布が、細胞表層における非対称な分布へと変化していくプロセスの経時観察を試みました。この実験のため、細胞内の金属濃度によって遺伝子の発現を開始する遺伝子プロモーター[7]を利用し、培養液に金属を添加することで任意のタイミングでPar3を発現させ、非対称性を作り出す実験システムを構築しました。この実験システムを利用した観察の結果、Par複合体の非対称な分布は、①点状のPar複合体の凝集体が細胞の表層に出現する、②点状のPar複合体の凝集体がより大きな島状の構造(Par-island)へ成長し、離散と集合を繰り返す、③島状の構造が細胞の一部に偏った状態でその分布が安定化する、という三つのステップを経て形成されていくことが明らかになりました。(図2)
また、Par複合体システムによる極性を持つ典型例であるショウジョウバエの神経幹細胞を観察したところ、Par-islandと同様の構造が観察されました。これは、Par-islandは培養細胞系だけの人工的な産物ではなく、Par極性が形成される際に一般的に形成される構造であることを示しています。このPar-islandは、不定形で動的な島状の構造を持つだけでなく、常に離散と集合を繰り返すというユニークな特徴を持った凝集体です。このことは、Par複合体は細胞内において液体状の性質を示し、凝集する相分離の状態にあることを示唆しています。
次に、Par-islandの構造や形成メカニズムをより詳しく検証するため、超解像顕微鏡による観察を行ったところ、Par-islandには網目状の構造が存在することが分かりました(図3)。この網目構造を構成する「ひも」の長さを計測すると、約400ナノメートル(nm、1nmは10億分の1メートル)の比較的均一なセグメントの組み合わせから成り立っていることが観測されました。これは、Par-islandの網目構造にはPar複合体からなるセグメント状の構築単位があることを示しています。
さらに詳しい動態の観察から、Par-islandが常に変形するという特徴は、(1) 個別のヒモ構造あるいは少数のヒモの集合体が、常時Par-islandに対して結合と解離を繰り返していること、(2) 網目構造の柔軟な組み換えがPar-island内で起きていること、という二つの要因によって引き起こされていると考えられます。
今後の期待
本研究では、極性を持たない培養細胞で人為的に細胞の非対称性を作り出すことで、細胞極性を再構成することに成功しました。そして、この再構成実験系を利用することで、Parタンパク質複合体による極性形成の基本的なプロセスや、その際に観察されるユニークな凝集体の微細な構造およびその動態を明らかにしました。
本研究で明らかになった極性形成のメカニズムをさらに発展させ、Parタンパク質複合体のセグメント構造の形成位置を生体内で自由に制御できれば、細胞極性の向きや形成タイミングを人為的に操作する手法の開発が期待できます。これにより、組織の中の細胞の配置を人為的に操作し、より自由度の高い器官再生が実現する可能性があります。
本研究の基盤となっている人為的な細胞極性の再構成システムは、薬剤添加などの処理に対する細胞極性の応答の観察などに応用できます。本システムは、細胞極性に関する分子生物学的な研究の推進だけでなく、細胞極性の乱れによって引き起こされるがんなど、さまざまな異常や疾患の研究など幅広く利用できると考えられます。
原論文情報
Kalyn Kono, Shigeki Yoshiura, Ikumi Fujita, Yasushi Okada, Tatsuo Shibata, Fumio Matsuzaki, “Reconstruction of Par-dependent polarity in apolar cells reveals a dynamic process of cortical polarization”, eLife, 10.7554/eLife.45559.001
発表者
理化学研究所
生命機能科学研究センター 非対称細胞分裂研究チーム
チームリーダー 松崎 文雄(まつざき ふみお)
(京都大学大学院生命科学研究科 教授)
大学院生リサーチ・アソシエイト 河野 夏鈴(こうの かりん)
(京都大学大学院生命科学研究科 博士課程)
お問い合わせ先
山岸 敦(やまぎし あつし)
報道担当
理化学研究所 広報室 報道担当
京都大学総務部広報課 国際広報室
補足説明
-
- 細胞極性
- 細胞を構成する細胞膜や細胞内の成分が非対称的に分布することで形成される、細胞の空間的な方向性(異方性)。移動する細胞の方向決定、細胞集団に境界を作る働き、あるいは、細胞極性の方向を細胞分裂の方向を一致させることにより、異なる娘細胞を生み出す非対称細胞分裂などのさまざまな基本的な細胞機能の過程に重要な役割を果たす細胞の特性。
-
- Parタンパク質
- 元々は線虫の非対称分裂に関わる遺伝子として見つかった、par-1からpar-6の6種類の遺伝子から産生されるタンパク質をさしていた。その後、ショウジョウバエの既知遺伝子との相同性があることや、哺乳類で相同な遺伝子が発見されたことなどから、種を超えて保存された機能があることが分かった。特に、Par3、Par6とaPKC(非定型プロテインキナーゼC)は複合体を形成し、細胞の非対称性の成立に中心的な役割を果たすサブセットの一つである。
-
- 超解像顕微鏡
- 従来の光学顕微鏡とは異なる原理を用いて、回折限界によって決まる分解能の限界(約200nm)よりも細かい対象物を解像できる光学顕微鏡。
-
- 神経幹細胞
- 自己複製能と多分化能の両方を持つ、ニューロンなど神経系の組織幹細胞。
-
- 上皮細胞
- 細胞に表裏があり、その一方に隣接細胞と接着する構造があり、他方は基底膜に接しする。互いに同じ方向を向いて接着することにより、細胞の集団に境界を引く。例えば、体表面を覆ったり、消化管や気管など管腔臓器の粘膜を構成したりする。外気や液体などから内部構造を守る役割を持つ。
-
- 反応拡散系モデル
- 空間や液体中に分布する物質が、局所的な「反応」と自由な「拡散」という二つの性質を合わせ持つ場合、その物質がさまざまな空間的な濃度分布、時間的な変化、あるいは定常状態を示すことがアラン・チューリングによって提示され、生物の形態形成のモデルとして適していることが指摘された(1952年)。この理論を利用した生命現象の数理モデルを反応拡散系モデルという。実際、生物が示す時間的、あるいは空間的なパターンの多くをよく説明し、形態形成の一つの一般的な原理とされている。
-
- プロモーター
- ゲノムDNA上で遺伝子(RNA)として転写される領域の近くにあり、遺伝子を発現させる機能を持つ領域(配列)。
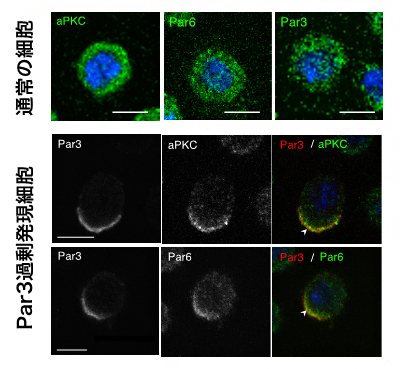
図1 培養細胞において Par3の発現上昇により生じたタンパク質の非対称分布
上段) 通常の状態では、Par複合体を構成するaPKC、Par6、Par3の各タンパク質は、細胞質に均一に分布している(緑色は各タンパク質の分布を、青色は細胞核を示す)。スケールバーは5μm。
中下段) Par3の発現量を人為的に上昇させた細胞では、Par3が細胞表層で非対称な分布に変化するとともに(左)、aPKCとPar6の分布も同様な非対称性を示した(中)。このとき、各タンパク質の局在は一致していたことから(右、矢頭)、Par複合体の非対称分布によるPar極性の出現が確認された。スケールバーは5μm。
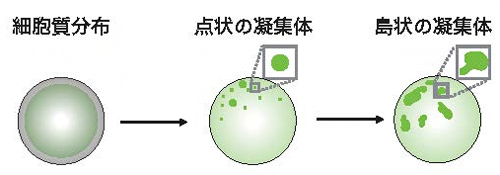
図2 培養細胞上で非対称性が生まれる過程の概略図
操作前では細胞質内に均一に分布しているPar複合体は、まず点状の凝集体として細胞表層に分布する。時間の経過とともに、点状の凝集体は島状の凝集体(Par-island)へと成長する。この凝集体は離散と集合を繰り返しながら、細胞表層の一部へと集合する。その結果、細胞内でタンパク質の非対称な分布が形成される。
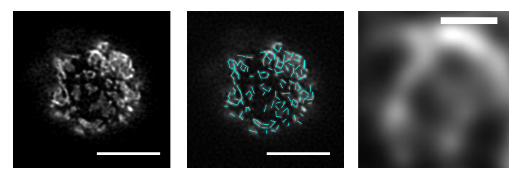
図3 Par-islandに内包されていた網目構造
Par-islandを超解像顕微鏡によって観察した様子。中央は、画像からセグメント構造をトレースしたもの。400ナノメートル程度のセグメント構造が集合し、網目構造を形成することによって、Par-islandの島状の構造が形成されていることが分かる。スケールバーは5μm(左、中央)、1μm(右)。

