琵琶湖産ボルボックスの全ゲノム情報から解明
2021-05-18 東京大学
山本 荷葉子(生物科学専攻 博士課程3年生(研究当時))
野崎 久義(大学院理学系研究科 特任研究員/生物科学専攻 准教授(研究当時))
発表のポイント
- 琵琶湖産ボルボックス(注1)の雌雄異体種(メスとオスが異なる種)と雌雄同体種の全ゲノム比較解析から、雌雄同体種は、雌雄異体種のメス側の性染色体が残存したままオス特異的遺伝子を獲得することで誕生したことが明らかになった。
- 雌雄異体種と雌雄同体種との転換進化はさまざまな生物で研究されているが、雌雄同体種への転換時におけるメスとオスの性染色体の運命を初めて明らかにした。
- 雌雄同体種へ進化した後もメス側の性染色体だけがほぼ残存している驚くべき事実はメス機能とオス機能の発現の差異を反映している可能性があり、性の進化の新たなる研究が期待される。
発表概要
神話や伝説では雌雄同体の神が世界各地で知られていますが、無脊椎動物や植物・藻類では実際に雌雄同体(同株)の種が進化しており、古くから進化生物学者の興味を惹いています。しかし、メスとオスで異なる種類の性染色体(注2)をもつ種(雌雄異体種)からどのようにして両性の特徴を同時に持つ生物が誕生したかは謎に包まれていました。
今回、東京大学大学院理学系研究科と国立遺伝学研究所等の研究グループは琵琶湖産のボルボックス属に属する互いに近縁な2種(図1, 2)の次世代シーケンス(注3)を用いた全ゲノム解読を実施し、雌雄同体種が誕生した直前と直後に相当する生物の性染色体の全貌を明らかにしました(図3, 4)。
その結果、祖先種のメス側の性染色体が残存したまま”OTOKOGI” (注4)等のオス特異的遺伝子を獲得することで雌雄同体種が誕生したことが明らかになりました。雌雄同体種へ進化した後もメス側の性染色体だけがほぼ残存しているという驚くべき事実は、有性生殖におけるメスとオスの根源的な差異を反映している可能性があり、性の進化研究の新たなる展開が期待されます。
発表内容
研究の背景
私たちヒトのような哺乳類はメスとオスが別個体で、両性で異なる性染色体をもっています(雌雄異体種)。一方、無脊椎動物や植物・藻類のさまざま系統では一つの個体がメスとオスの両方の配偶子をつくる雌雄同体(同株)の種が進化しています。自家受精して遺伝的多様性を低下させる可能性のある雌雄同体種の進化は古くから進化生物学の大きな問題であり(文献1)、現在でも活発に研究が展開されています。しかし、哺乳類のようなXY型性染色体(注5)をもつ生物群では雌雄同体種への進化が認められないことと、比較生物学的研究に適当な雌雄異体種と近縁な雌雄同体種を材料とすることが困難であったため、雌雄同体の新種誕生の過程で祖先種のメスとオスの性染色体がどのような運命を辿るかは不明でした。
研究内容
今回、東京大学大学院理学系研究科生物科学専攻の野崎久義准教授(研究当時)のグループはこの進化を実験的に解明できる生物として、琵琶湖(図1)から自ら採集した緑藻ボルボックス属に属する互いに近縁な2種(文献2)を研究に用いました(図2)。野崎のグループは以前から、Starr(1971)が世界各地から採集しボルボックス・アフリカヌス(学名:Volvox africanus) と同定した培養株が4タイプの多様な有性生殖を持つこと(文献3)に注目していました。しかし、Starr (1971)の培養株はほとんど死滅しており、研究に使用できない状態でした。このためには新たなる培養株をフィールド調査から得る必要があると考えていました。その結果、野崎のグループは琵琶湖からStarr (1971) の2タイプに相当する新規培養株を確立することに成功しました(文献2)。これら2タイプは種としても異なることが判明し、ボルボックス・アフリカヌスと新種ボルボックス・レティクリフェルス(学名:V. reticuliferus)に分類しています(文献2)。両種は約1千万年前に分岐し、ボルボックス・アフリカヌスはメスとオスの両方の配偶子が同一の培養株で形成される雌雄同体(同株)種であり、ボルボックス・レティクリフェルスはメスとオスが遺伝的に異なる雌雄異株種であります(図2)。

図1:日本の古代湖「琵琶湖」におけるボルボックス類の現地調査。琵琶湖には日本の他の場所では見ることのできないボルボックスが2種(図2)生育する。これら2種が今回の全ゲノム解析に用いられた。野崎撮影。
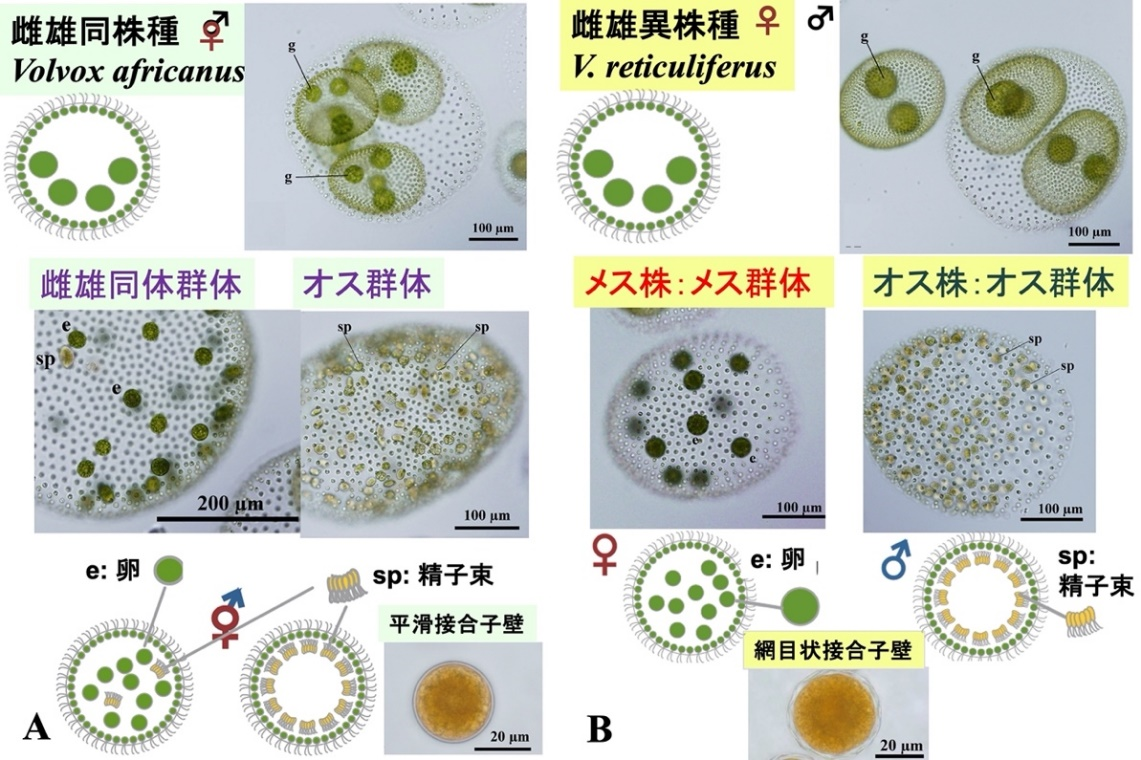
図2:今回の全ゲノム解析に用いた琵琶湖産のボルボックス2種。A.雌雄同株種ボルボックス・アフリカヌス。 B.雌雄異株種ボルボックス・レティクリフェルス。今回の成果と文献2の図を改変。
今回はボルボックス・アフリカヌス並びにボルボックス・レティクリフェルスのメス・オス、合計3生物の次世代シーケンスを用いた全ゲノム解読を実施し、雌雄同体種が誕生した直前と直後に相当する生物の性染色体の全貌を明らかにしました(図3)。雌雄異株種ボルボックス・レティクリフェルスの性染色体上のメスとオスで配列の異なる性決定領域(SDR)は約100万塩基対と非常に大きく、オスのSDRには “OTOKOGI” 等のオス特異的遺伝子が3個、メスのSDRにはメス特異的遺伝子が3個位置していました。雌雄同株種ボルボックス・アフリカヌスでもSDR に非常に類似した約100万塩基対の性決定類似領域(SDLR)が1個保持されており、系統解析から雌雄異株種のメスのSDRに由来することが明らかになりました。一方、オスのSDRに由来すると考えられる“OTOKOGI” 等のオスの特性に重要な遺伝子は、SDLR 以外のゲノム上の離れた場所に存在していました(図3)。また、雌雄同株種では ”OTOKOGI” が5個に縦列重複してクラスターを形成するという非常に興味深い結果を得ました(図3)。
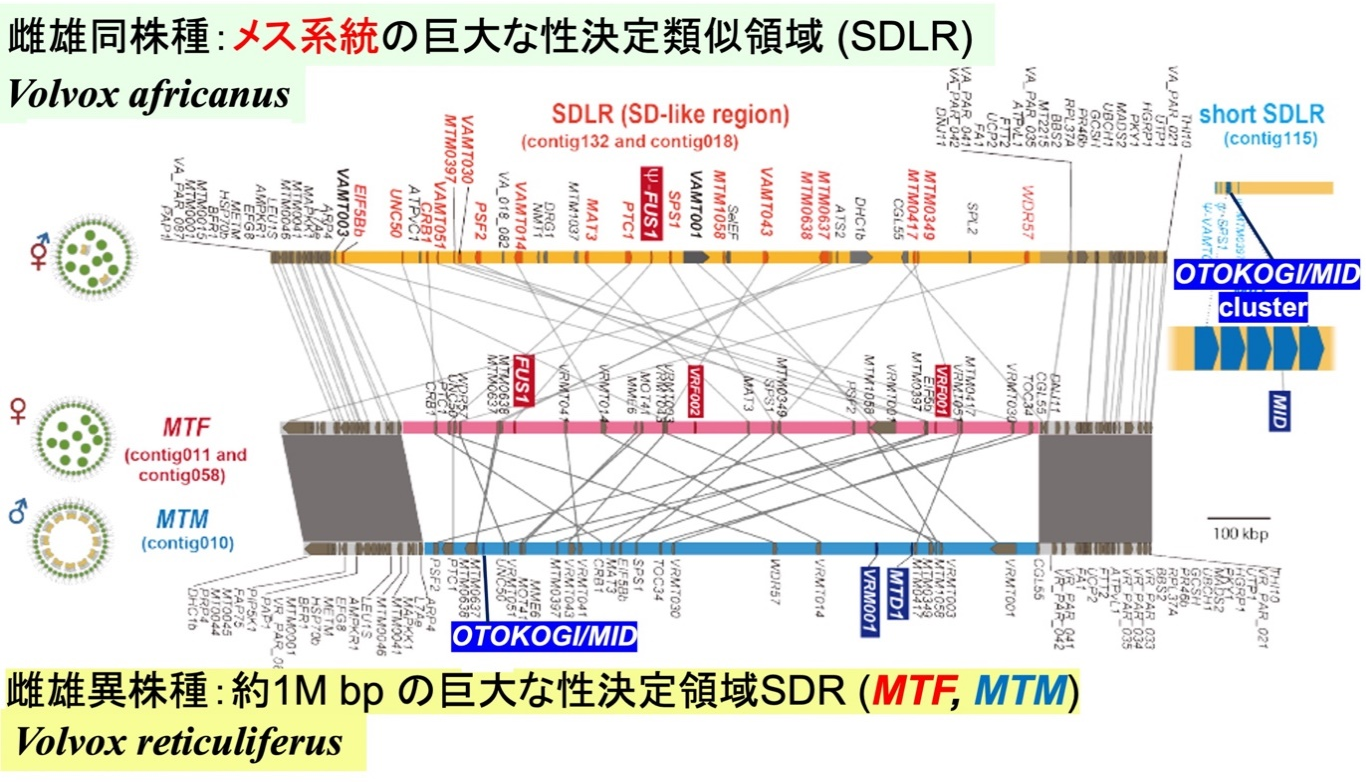
図3:全ゲノム解析の結果明らかになったボルボックス2種(図2)の性染色体上の性決定領域(SDR)と性決定類似領域(SDLR)。今回の成果に基づく。
ボルボックス・レティクリフェルスが約7千5百万年前に分岐したボルボックス・カルテリ(Volvox carteri)と同様に(文献4)約100万塩基対と非常に大きなSDRを性染色体上に保持していたことは、巨大なSDRがボルボックスの有性生殖で大きな意義を持つことを示唆します(図4)。さらに、雌雄同株種に進化しても祖先種のメスの性染色体のSDRが残存して、重要なオスの遺伝子がバラバラに存在することは、祖先種のメス側の性染色体を残存させたままオスの機能に重要な ”OTOKOGI” 等の遺伝子を獲得して、メスとオスの両方の機能を備える雌雄同株種が誕生したことを意味します。ボルボックス系列(注6)において ”OTOKOGI” (MID) は、異型配偶・卵生殖の生物のオスまたは同型配偶(注7)生物のマイナス交配型に必ず存在する遺伝子であり(文献5)(図4)、同型配偶の生物ではマイナス交配型を決定します(文献6)。最近の研究からボルボックス・カルテリのメスに“OTOKOGI”遺伝子を導入すると精子を形成するようになることが明らかになっており(文献7)、この遺伝子単独でメスの中にオスの機能を付加できることを示しています。一方、メスの機能を決定する、またはメスに必須の遺伝子は知られていません(文献5)(図4)。
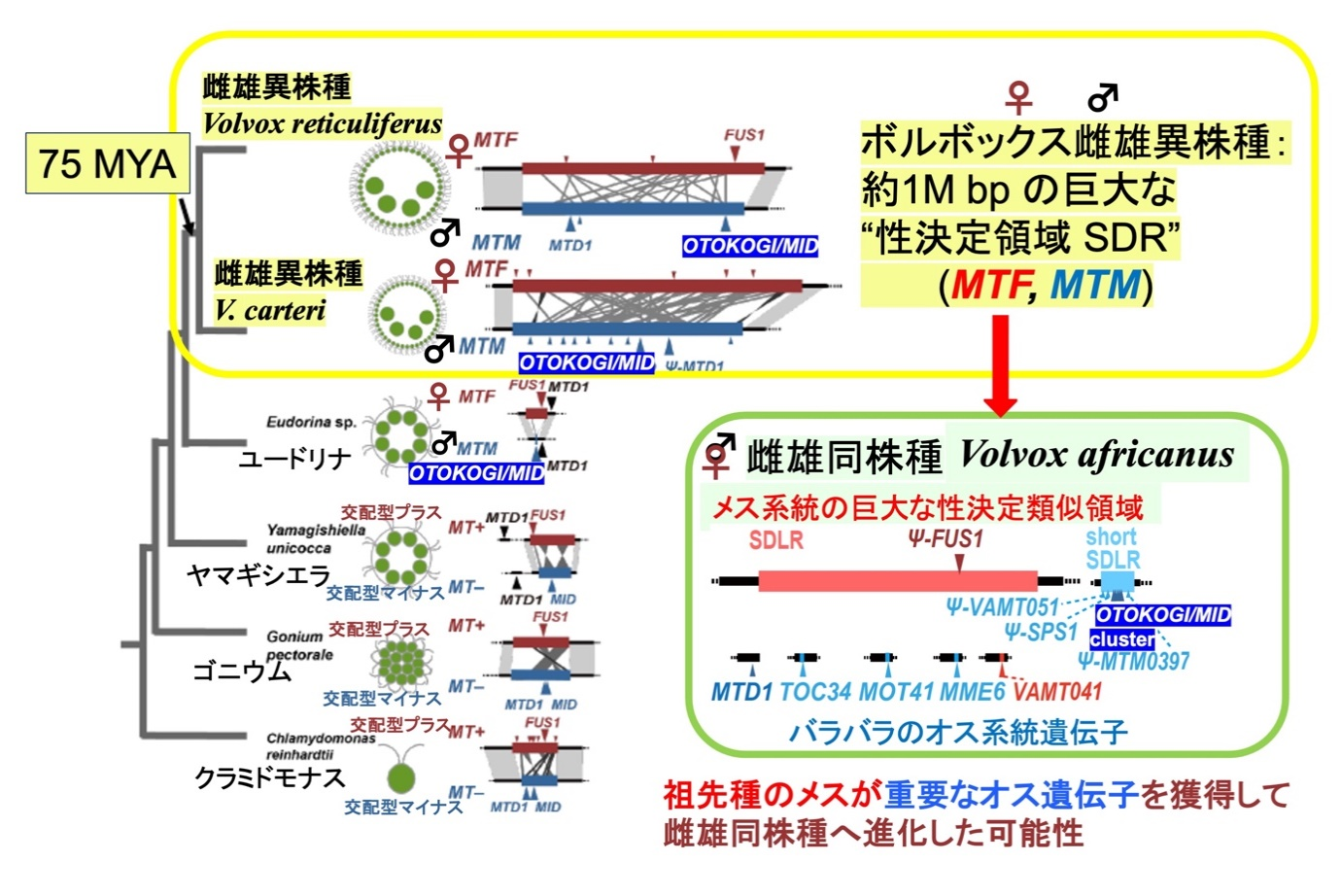
図4:緑藻ボルボックス系列の系統関係と性染色体の進化。今回の成果に基づく。
したがって、今回明らかになった雌雄同株種における祖先種のメスとオスの性染色体の残存状態の差異(図3, 4)は、メス機能とオス機能の発現機構の差異に関連する可能性を示唆しています。すなわち、メスの性染色体の巨大なSDRが総和としてボルボックスのメスの機能を担っている可能性が考えられます。
今後の予定
雌雄同株種へ進化した後もメス側の性染色体の巨大なSDRだけがほぼ残存しているという驚くべき事実はメスとオスの機能発現の有性生殖における本質的な差異を反映している可能性があり、性の進化研究の新たなる展開が期待されます。
本研究は第一著者山本荷葉子の令和2年度本理学系研究科の博士論文の一部であり、先進ゲノム支援等の文部科学省・日本学術振興会の科学研究費補助金(18J11391 山本荷葉子, 16H06279 豊田敦, 16H02518 野崎久義, 20H03299 野崎久義, 17K07510 豊岡博子, 17H05840 西村芳樹・浜地貴志)の支援を受けて行われました。
参考文献
1. C. Darwin, The correspondence of Charles Darwin. Vol. 9. F. Burkhardt, J. Browne, D. M. Porter, M. Richmond, Eds. (Cambridge University Press, 1861).
2. H. Nozaki, R. Matsuzaki, K. Yamamoto, M. Kawachi, F. Takahashi. Delineating a new heterothallic species of Volvox (Volvocaceae, Chlorophyceae) using new strains of ‘Volvox africanus’. PLoS ONE 10, e0142632 (2015).
3. R. C. Starr. Sexual reproduction in Volvox africanus. In: Parker BC, Brown RM Jr., editors. Kansas: Contribution in Phycology, Allen Press; 1971. pp. 59–66.
4. P. Ferris et al., Evolution of an expanded sex-determining locus in Volvox. Science 328, 351–354 (2010).
5. T. Hamaji et al., Anisogamy evolved with a reduced sex-determining region in volvocine green algae. Commun. Biol. 1, 17 (2018).
6. P. J. Ferris, U. W. Goodenough. Mating type in Chlamydomonas is specified by mid, the minus-dominance gene. Genetics 146, 859–869 (1997).
7. S. Geng et al., Evolution of sexes from an ancestral mating-type specification pathway. PLoS Biol. 12, e1001904 (2014).
発表雑誌
- 雑誌名
Proceedings of the National Academy of Sciences of the United States of America (PNAS)論文タイトル
Three genomes in the algal genus Volvox reveal the fate of a haploid sex-determining region after a transition to homothallism著者
Kayoko Yamamoto, Takashi Hamaji, Hiroko Kawai-Toyooka, Ryo Matsuzaki, Fumio Takahashi, Yoshiki Nishimura, Masanobu Kawachi, Hideki Noguchi, Yohei Minakuchi, James G. Umen, Atsushi Toyoda and Hisayoshi Nozaki*DOI番号
10.1073/pnas.2100712118アブストラクトURL
Just a moment...www.pnas.org

用語解説
注1 ボルボックス
淡水産緑藻類の一属 Volvoxで、世界各地から20種以上が報告されている。500以上の2鞭毛を持つ細胞が球体の表層に鞭毛を外側に向けて遊泳し、しばしば直径1 mmを超える。「群体性生物」として紹介される場合があるが、ボルボックスの「群体」には細胞分化もみられ、実態としては多細胞生物である(本文中ではこれまでの伝統的な名称「群体」を便宜的に使用している)。無性生殖と有性生殖の両方で増殖する。有性生殖では卵と精子を形成する。
注2 性染色体
メスとオスで個体が異なる生物における、雌雄で形や大きさが異なる染色体。もしくは性を決定するゲノム領域(性決定領域SDR)が存在する染色体。性染色体以外の、雌雄で共通な染色体を常染色体という。
注3 次世代シーケンス
革新的な装置である次世代シーケンサーで塩基配列を大量・高速・安価に配列決定すること。磁気ビーズ上に増幅したDNA断片を用いて配列決定する方法がこれまで用いられてきたが、この方法では長い配列は得られなかった。最近、塩基配列を1分子ずつ電気的に読みだす手法により非常に長い配列を解読する次世代シーケンサーが普及した。本研究ではこれらの両手法を組み合わせることで、それぞれの弱点を補い、長くて正確なゲノム解読が実現した。
注4 ”OTOKOGI”
同型配偶(注7)の緑藻クラミドモナスでは、交配型マイナスを決定する遺伝子としてMID 遺伝子が知られている(文献6)。オスとメスが分化したボルボックス系列のプレオドリナでは MID と起源を同じくする遺伝子 (PlestMID) がオスだけに存在し、“OTOKOGI”と呼ばれる。
注5 XY型性染色体
複相(2n)の生物において、オスがXとY(異型)、メスがXを2本持つ(同型)性決定様式で観察される性染色体。
注6 ボルボックス系列
淡水域に生育する2鞭毛をもつ単細胞生物クラミドモナスと細胞が4個以上で集合した多細胞生物から構成される緑藻の一群。各細胞には葉緑体が1個ずつ含まれ、光合成を行う。多細胞性の種は単細胞性のクラミドモナスのようなものから進化したと考えられており、500細胞以上からなり小さい非生殖細胞(分裂をして無性生殖しない)が分化してオス・メスをもつボルボックスと、オス・メスが未分化な同型配偶のクラミドモナスとの間に、ゴニウム、ヤマギシエラ、ユードリナなど体制と有性生殖の観点で進化的に中間段階の生物が現存するため、性の進化と多細胞化のモデル生物群と考えられている。図4参照。
注7 同型配偶
合体する配偶子に大きさと運動能力の差が認められない、雌雄が未分化な有性生殖の一形態。同型配偶の生物では配偶子の大きさ等に差がないため、異なる性(交配型)を便宜的に交配型マイナスまたはプラスとして区別している。雌雄が分化した有性生殖を異形配偶または卵生殖(一般的にメスの配偶子が鞭毛を欠き不動の場合)という。

