2022-10-02 東京大学
安藤 英伍(日本学術振興会特別研究員)
木下 俊則(名古屋大学トランスフォーマティブ生命分子研究所 教授)
寺島 一郎(生物科学専攻 教授)
発表のポイント
- 植物の気孔孔辺細胞の細胞膜H+ポンプが、二酸化炭素(CO2)濃度の上昇に応じて脱リン酸化されることで気孔閉鎖が促進される仕組みを明らかにしました。
- 光合成に用いられるCO2の取込み口である気孔が、CO2自体によって制御される新たな仕組みを提唱しました。
- 光合成活性が変動する野外での気孔開閉の仕組みの解明や大気CO2濃度の削減に向けた作物のCO2吸収の効率化技術の開発に繋がることが期待されます。
発表概要
植物の表面には気孔と呼ばれる孔(あな)が多数存在し、光合成に必要なCO2や蒸散(注1)に係る水の通路として機能していることから、気孔の開閉制御は植物の成長・生存に重要な役割を果たしていると考えられています。植物に光が当たると、気孔を形成する孔辺細胞の細胞膜H+ポンプ(注2)と呼ばれるタンパク質が、リン酸化(注3)によって活性化して気孔開口を駆動します。一方、暗黒下では気孔が閉鎖しますが、その仕組みは明確にされていませんでした。
今回、安藤英伍日本学術振興会特別研究員(東京大学大学院理学系研究科 生物科学専攻 植物生態学研究室所属)、名古屋大学トランスフォーマティブ生命分子研究所の木下俊則教授、東京大学大学院理学系研究科の寺島一郎教授らのグループは、暗黒下での光合成の停止に伴う葉内CO2濃度の上昇と並行してH+ポンプが急激に脱リン酸化される現象を発見しました。研究グループは葉内CO2濃度を人為的に上昇させることで、光照射下であってもH+ポンプの脱リン酸化を引き起こせることを明らかにしました。さらに遺伝学的検証から、欠損するとH+ポンプの脱リン酸化が損なわれ、気孔の閉鎖が遅延する遺伝子を同定しました。
本発見は、光合成に使われるCO2の取込み口である気孔が、CO2自体によって制御される新たな仕組みを提唱するものです。今後、光合成活性が変動する野外での気孔開閉の仕組みの解明、大気CO2濃度の削減に向けた作物のCO2吸収効率化技術への応用が期待されます。
発表内容
植物は光に応答して気孔を開口し、光合成に必要なCO2の取込みや蒸散とそれを通じた根からの水分や養分の吸収を行っています。このことから、気孔開閉は植物の成長・生存、さらには生態系における炭素・水循環に重要であるとして広く研究されてきました。太陽光中に含まれる青色光は孔辺細胞に受容され、細胞膜H+ポンプのリン酸化を通じた活性化を引き起こし、活性化したH+ポンプの作用により気孔開口が駆動されます。さらに近年、光合成もH+ポンプのリン酸化に寄与することが発見されました(図1)。一方、暗所では気孔が閉鎖しますが、その初期過程については明確にされていませんでした。

図1: 気孔の機能と光による気孔開口の仕組み
植物は光に応じて気孔を開き、光合成に必要なCO2の取り込みや、蒸散とそれを通じた根からの水分(H2O)および養分の吸収を促進する。太陽光中に含まれる青色光や光合成を介して、孔辺細胞の細胞膜H+ポンプの活性化に重要なリン酸化が引き起こされる。活性化したH+ポンプの働きにより、気孔開口に必要な細胞内へのカリウムイオン(K+)などの蓄積が駆動される。
今回、本研究グループは、モデル植物シロイヌナズナを用いて、葉を光照射下から暗黒中に移動、すなわち光合成が停止する際の孔辺細胞のH+ポンプのリン酸化状態を蛍光により可視化して解析しました(図2A)。すると、驚いたことに暗黒に移してわずか1分でH+ポンプが脱リン酸化されることを発見しました(図2B)。この現象が光合成の停止による葉内CO2濃度の上昇と相関することから、研究グループは人為的に葉内CO2濃度を上昇させてもH+ポンプの脱リン酸化を引き起こせるのではないかと仮説を立て検証しました。その結果、光照射下であっても葉内CO2濃度を2倍程度に増やすと脱リン酸化が起こることが明らかになりました(図2C)。このことは、CO2がH+ポンプの脱リン酸化引き起こしていることを示しています。

図2: 暗黒やCO2に応答したH+ポンプの急速な脱リン酸化の発見
(A)孔辺細胞におけるH+ポンプのリン酸化を、抗体を用いて蛍光により可視化した。
(B)光照射下から暗黒への移行後1分でリン酸化レベルが低下 (脱リン酸化)することが示された。(上)蛍光顕微鏡画像 矢尻: 気孔孔辺細胞 スケールバー: 50 μm (下)孔辺細胞における蛍光強度(リン酸化レベル)の定量結果
次に研究グループは、CO2によってH+ポンプが脱リン酸化される仕組みの解明に取り組みました。細胞内に入ったCO2はカルボニックアンヒドラーゼ(炭酸脱水酵素; CA)と呼ばれる酵素(注4)の働きによりHCO3–に変換され、それが細胞内でシグナルとして作用すると考えられています。そこでカルボニックアンヒドラーゼをコードする遺伝子CA1とCA4を欠損する変異体(注5)植物を調べたところ、H+ポンプのすばやい脱リン酸化が損なわれており、気孔の閉鎖が遅くなることが示されました(図3A)。このことはカルボニックアンヒドラーゼを介した細胞内シグナル伝達(注6)によってH+ポンプの脱リン酸化、その後の気孔閉鎖が制御されていることを示しています。さらに、H+ポンプの脱リン酸化を触媒する酵素であることが最近示されたグループDの2C型プロテインホスファターゼをコードする遺伝子PP2C.D6とPP2C.D9を欠損する植物においてもH+ポンプの脱リン酸化が損なわれ、気孔閉鎖が遅延したことから(図3B)、CO2シグナル伝達の下流でこれらの脱リン酸化酵素が機能していることが示唆されました。
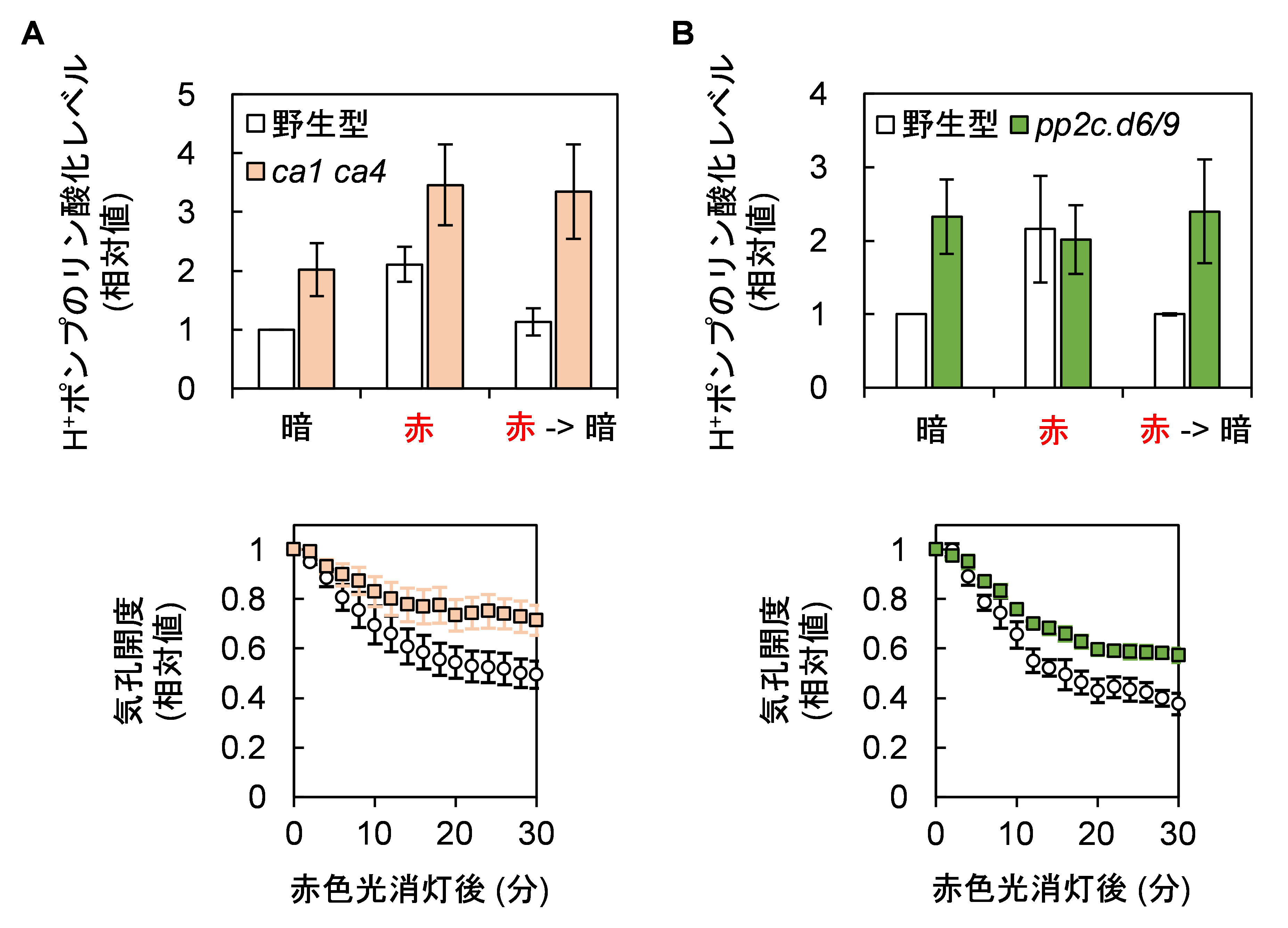
図3:H+ポンプの脱リン酸化に関与する因子
細胞内でCO2からHCO3–への変換を触媒する酵素であるカルボニックアンヒドラーゼを欠損する変異体(ca1 ca4; A)や H+ポンプの脱リン酸化を触媒するグループDの2C型プロテインホスファターゼを欠損する変異体 (pp2c.d6/9; B)では、暗所への移行によるH+ポンプの急速な脱リン酸化が起こらず (上)、素早い気孔の閉鎖も緩慢になった(下)。
以上の結果から、植物は光合成の停止をCO2濃度の上昇という形で検知しており、気孔開口の駆動力を形成するH+ポンプをPP2C.Dを介して直ちに脱リン酸化・不活性化して、効率よく気孔を閉じられるようにしていると考えられます(図4)。

図4:H+ポンプの脱リン酸化を介した気孔閉鎖のモデル
暗黒下での光合成の停止や環境中のCO2濃度の上昇により、葉内CO2濃度が上昇すると、カルボニックアンヒドラーゼ(CA)を介してグループDの2C 型プロテインホスファターゼ(PP2C.D)が働き、H+ポンプの脱リン酸化を通じて気孔開口の仕組みを抑制することで気孔の閉鎖が促進されると考えられる。
実は文献を調査すると、CO2がH+ポンプの働きを阻害する可能性は、1985年に示唆されていたのですが(Edwards & Bowling, Journal of Experimental Botany, 1985)、その仕組みは不明なままでした。本研究成果は、この長らく不明であったCO2によるH+ポンプの阻害の仕組みを30余年越しに明らかにするものでもあります。また、本研究では興味深いことに、光照射下にあってもCO2濃度の上昇はH+ポンプを脱リン酸化させる作用を示すことが分かりました。今後、光合成活性が変動する日中の野外環境での気孔開閉の仕組みの解明に繋がると考えられます。さらに大気CO2濃度削減に向けて、作物のCO2応答性を制御し、CO2吸収効率を向上させる技術の開発への応用も期待されます。本研究は特別研究員奨励費(課題番号: 20J00392)および 科研費(課題番号: 20H05687 および 20H05910)の支援により実施されました。
発表雑誌
- 雑誌名
New Phytologist論文タイトル
Elevated CO2 induces rapid dephosphorylation of plasma membrane H+-ATPase in guard cells
著者
Eigo Ando*, Hannes Kollist, Kohei Fukatsu, Toshinori Kinoshita*, Ichiro Terashima
DOI番号 10.1111/nph.18472
用語解説
注1 蒸散
水が主に気孔を通じて水蒸気として植物体内から大気中に拡散していく現象。
注2 細胞膜H+ポンプ
細胞内のエネルギー分子であるアデノシン三リン酸(ATP)を消費して、細胞の外へ水素イオン (H+; プロトン)を排出するタンパク質。
注3 リン酸化・脱リン酸化
生物内のタンパク質などの有機物にリン酸基が付加される化学反応をリン酸化、その逆を脱リン酸化と呼ぶ。タンパク質の場合、その機能調節(活性化・不活性化)などに重要な役割を持つ。
注4 酵素
細胞内のさまざまな化学反応を触媒(反応を促進)するタンパク質の総称。
注5 変異体
細胞の持つ遺伝物質に変異(欠損など)が生じた個体をその遺伝子の変異体と呼ぶ。
注6 細胞内シグナル伝達
シグナル分子を受容した細胞内で順次起こるタンパク質の機能変化などを介した情報伝達の仕組み。
")
