
2022-09-16 東京大学医科学研究所
発表のポイント
- 遺伝暗号解読活性を失ったリボソームが翻訳中に単独で停滞し、Fap1がリボソーム両側のmRNAの動きの停止を感知してリボソームに分解の目印をつけることを明らかにしました。
- 既知の停滞目印であるリボソーム間の衝突を伴わずに停滞したリボソームにFap1が特異的に結合することを見出し、単独のリボソームに特化した停滞センサーを世界で初めて同定しました。
- 本成果は、単独のリボソームによる翻訳が盛んな神経細胞等における翻訳異常の研究に貢献し、リボソームの機能欠損に起因する疾患発症機構の更なる理解につながると期待されます。
発表概要
リボソーム(注1)は、mRNA(注2)上の遺伝暗号を解読しタンパク質を合成する翻訳活動を担います。翻訳中のリボソームの停滞はタンパク質合成の異常を示す危険信号であり、翻訳品質管理機構(注3)により認識・解消されます。多くの品質管理機構がリボソーム間の衝突を停滞の目印とする一方、衝突を伴わない単独で停滞したリボソームを対象とする停滞解消の仕組みは不明でした。
今回、東京大学 医科学研究所RNA制御学分野/大学院理学系研究科生物科学専攻の稲田利文教授と東北大学大学院薬学研究科の李思涵大学院生、ミュンヘン大学Gene CenterのRoland Beckmann教授らのグループは、単独のリボソームに特化した停滞センサー因子として出芽酵母のFap1を世界で初めて同定しました。Fap1が単独で停滞した機能欠損リボソームに目印をつけ、停滞の解消と共にリボソーム自身の分解を誘導することを明らかにしました。
リボソームプロファイリング法(注4)によるmRNA上のリボソーム結合領域の網羅的解析およびクライオ電子顕微鏡(注5)を用いた構造解析より、Fap1はmRNA上に単独に存在するリボソームを好んで結合し、両側のmRNAの動きの停止を感知することを見出しました。本成果は、単独のリボソームによる翻訳が盛んな神経細胞等におけるリボソームの機能異常と疾患発症との関連の更なる理解につながると期待されます。
本研究は、CREST(課題番号:JP 19gm1110010、研究代表者:稲田利文)、日本学術振興会科学研究費助成事業(課題番号:JP18H03977, JP19H05281、稲田利文;21H00267, 21H05710, 22H02606、松尾芳隆)、科学技術振興機構(JST)さきがけ(課題番号:JPMJPR21EE、研究代表者:松尾芳隆)、日本学術振興会特別研究員奨励費(課題番号:20J20445、李思涵)、海外特別研究員(池内健)などの支援を受けて行われました。
本研究成果は、米国科学誌「Molecular Cell」(9月15日付けオンライン版)に公表されました。
発表内容
研究の背景
細胞内でリボソームがmRNA上の遺伝暗号を解読しタンパク質を合成する翻訳活動を担います。翻訳中のリボソームの停滞は、タンパク質合成効率の低下や遺伝子産物の機能欠損等を示す危険信号であり、細胞が保持する翻訳品質管理機構(以下「品質管理」)により認識・解消されることで遺伝子発現(注6)の正確性が維持されます。
単一のmRNA分子に複数のリボソームが付着することが多いため、先頭のリボソームが強く停滞すると後続のリボソームが追いつき、リボソーム同士の衝突が起こります。リボソーム衝突はリボソーム停滞の目印と見なされ、合成途上の不完全なタンパク質の分解系RQC (Ribosome-associated Quality Control)をはじめとする多くの品質管理を誘導することが近年明らかになりました。出芽酵母におけるRQCの活性化は、リボソーム衝突を感知するセンサー因子であるHel2が触媒するリボソームタンパク質uS10のユビキチン化(注7)を引き金とすることを稲田教授らのグループが先駆けて報告してきました。
一方、リボソーム間の衝突を伴わずに単独で停滞したリボソームを対象とする停滞認識・解消機構は不明でした。従来のリボソーム停滞に関する研究では、衝突を伴わない停滞を効率よく誘導および追跡することが困難であり、単独のリボソームに特化した停滞センサーは未同定でした。
研究内容
本研究ではまず、リボソームの活性中心への変異導入が、出芽酵母における単独のリボソーム停滞の効率的な誘導と解析に有用であることを発見しました。リボソームプロファイリング法を用いた解析により、リボソームの小サブユニット中の18S rRNAの変異によって遺伝暗号解読活性を失ったリボソーム(以下「機能欠損リボソーム」)がmRNA上の翻訳伸長(注8)の開始点である開始コドンで単独で停滞することが明らかになりました(図1a)。
興味深いことに、機能欠損リボソームは18S NRD (18S Non-functional rRNA Decay)の基質として知られています。18S NRDは、リボソームタンパク質uS3のユビキチン化を目印に、機能欠損リボソームのサブユニットへの解離を経て18S rRNAを迅速に排除するリボソーム自身の分解系です。研究グループは、遺伝学的スクリーニングおよび生化学的な実験により、18S NRDにおけるuS3のユビキチン化はMag2とFap1という二つの因子が順次に触媒する二段階の反応であることを見出しました(図1b)。

図1. Fap1は単独で停滞した機能欠損リボソームに起因する品質管理に必須である
リボソームプロファイリングの結果から、機能欠損リボソームが開始コドンで停滞することがわかった。開始コドンでリボソームの大、小サブユニットが会合するため、その上流に翻訳伸長中のリボソームがなく、リボソーム同士の衝突が起きないと想定される。 18S NRDの模式図。機能欠損リボソームはuS3がMag2とFap1により二段階でユビキチン化された後サブユニットへ解離し、小サブユニット中の18S rRNAがエキソヌクレアーゼによって分解される。
これらの因子が認識するリボソームの特徴をリボソームプロファイリング法で解析した結果、Mag2が翻訳速度の遅いリボソームに全般的に結合する可能性が示唆された一方、Fap1は隣接リボソームの出現頻度の低い位置に存在するリボソームを好んで結合する傾向が見られました(図2)。また、Fap1の働きを補助する可能性のある因子としてYil161wおよびFpr1を質量分析(注9)により同定しました。
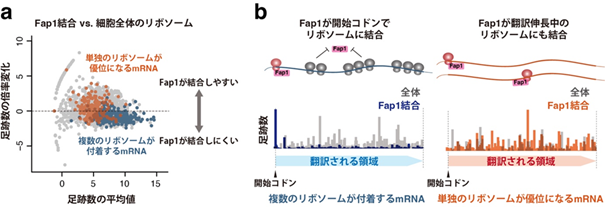
図2. Fap1は単独に存在する確率の高いリボソームを好んで結合する
Fap1が結合したリボソームと細胞全体のリボソームの足跡数(サンプル間の総足跡数を補正後)との比較。複数のリボソームが付着するmRNA(青色)とリボソームが一つのみ存在する頻度の高いmRNA(褐色)をハイライトしている。 それぞれのタイプのmRNAの代表例におけるFap1結合型リボソームの分布。
さらに、クライオ電子顕微鏡を用いた解析により、Fap1-Yil161w-Fpr1 (FAP複合体)がリボソームに結合している立体構造を捉えることに成功しました。Fap1は複数の亜鉛結合部位からなる細長い構造を介してリボソーム小サブユニットの頭部と呼ばれる部分にアーチをかけ(図3a)、両端のRINGドメインとR3Hドメインがリボソーム上のmRNAの入口、出口それぞれと相互作用する様子が観察されました(図3b-d)。翻訳伸長中のリボソームの構造変化によるmRNAの相対的な動きがこれらの相互作用を破壊し、リボソームが静止状態になった時のみFAP複合体が活性化する可能性が示唆されました。加えて、Fap1の結合部位と既知のリボソーム同士の衝突面との比較から、後続・衝突したリボソームの存在が停滞したリボソームへのFap1の結合を立体的に妨害することが分かりました(図3e)。
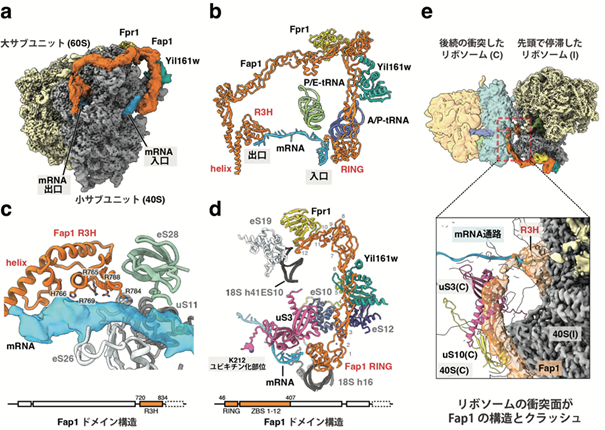
図3. Fap1はリボソームの両側のmRNAの状態を監視する
Fap1-Yil161w-Fpr1(FAP)とリボソームとの複合体のクライオ電子顕微鏡構造。 mRNAにFAPが結合した構造モデルの全体像。 mRNAの出口付近におけるFap1とmRNAとの相互作用に注目した構造モデルの拡大図。 mRNAの入口付近に注目した構造モデルの拡大図。 FAPが結合したリボソームと既知の衝突リボソームの構造との重ね合わせ図。
これらの結果から、Fap1はリボソーム両側のmRNAの動きの停止を感知することで、衝突を伴わない停滞をリボソームの機能異常の信号として提示し、サブユニット解離に伴う停滞の解消およびリボソーム自身の分解を誘導することが明らかになりました(図4)。
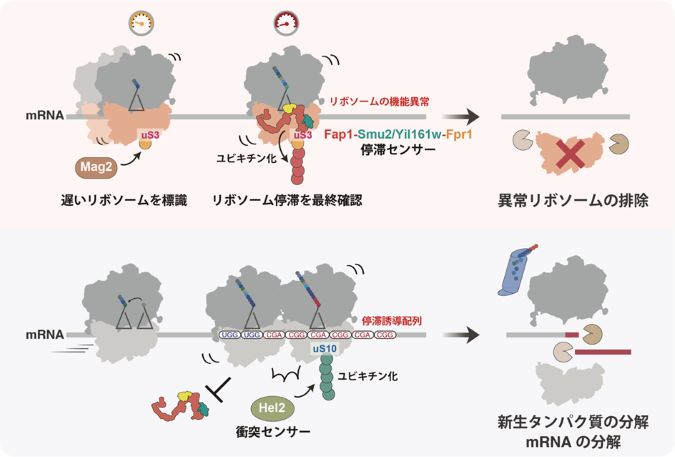
図4. 単独のリボソーム停滞とリボソーム衝突との比較
単独で停滞したリボソームが分解へと導かれる経路の模式図(上図)。Mag2が翻訳速度の遅いリボソームのuS3に一つ目のユビキチンを付加後、リボソームが単独で停滞した場合、停滞センサー因子Fap1が結合しユビキチン鎖を伸長させる。uS3に付加されたユビキチン鎖がリボソームの機能異常の目印と見なされ、リボソーム自身の分解・排除を引き起こす。一方、停滞したリボソームに後続のリボソームが衝突した場合(下図)、Fap1の結合が妨害され、衝突センサー因子Hel2によるuS10のユビキチン化およびそれに依存した品質管理が活性化する。
社会的意義・今後の予定
正確なタンパク質合成は生命活動の基盤であり、翻訳の異常はさまざまな疾患の原因になります。ヒトにはFap1をはじめとする18S NRD関連因子が保存されており、出芽酵母に類似した翻訳異常に対応するための品質管理が備わっていることが想定されます。衝突を伴わないリボソーム停滞の感知・解消機構を解析した本研究の成果は、単独のリボソームが主役となる神経突起での局所的翻訳等における品質管理研究の切り口になり、リボソームの機能異常および品質管理の破綻と疾患発症との関連の更なる理解にもつながると期待されます。
発表雑誌
雑誌名:「Molecular Cell」(9月15日付けオンライン版)
論文タイトル:Sensing of individual stalled 80S ribosomes by Fap1 for non-functional rRNA turnover
著者:Sihan Li†, Ken Ikeuchi†, Misaki Kato, Robert Buschauer, Takato Sugiyama, Shungo Adachi, Hideo Kusano, Tohru Natsume, Otto Berninghausen, Yoshitaka Matsuo, Thomas Becker, Roland Beckmann*, Toshifumi Inada*
†共同第一著者
*共同責任著者
DOI:doi.org/10.1016/j.molcel.2022.08.018
URL:http://doi.org/10.1016/j.molcel.2022.08.018
問い合わせ先
<研究に関するお問い合わせ>
東京大学医科学研究所 RNA制御学分野
教授 稲田 利文(いなだ としふみ)
<報道に関するお問い合わせ>
東京大学医科学研究所 国際学術連携室(広報)
用語解説
(注1)リボソーム
リボソームタンパク質とリボソームRNA (rRNA)から構成される巨大な複合体であり、mRNAにコードされている遺伝暗号(コドン)に従ってアミノ酸同士を結合させ、タンパク質を合成する装置。全体として大小二つのサブユニットで構成されている。
(注2)mRNA
メッセンジャーRNAの略で日本語では伝令RNA。タンパク質合成の設計図となる遺伝情報を持つRNA。
(注3)翻訳品質管理機構
異常な遺伝子産物を認識し分解することで、遺伝子発現の正確性を保証するシステム。
(注4)リボソームプロファイリング法
細胞からリボソームを抽出し、リボソームが覆っているmRNAの配列を次世代シーケンシングにより網羅的に同定、定量する解析法。
(注5)クライオ電子顕微鏡
液体窒素(-196℃)等により極低温に冷却された試料に対して電子線を照射し、試料を透過した電子線を検出することにより試料の観察を行う顕微鏡。
(注6)遺伝子発現
遺伝子の情報が生体機能をもつタンパク質などに変換されることを指す。
(注7)ユビキチン化
ユビキチンは76アミノ酸からなる低分子タンパク質である。ユビキチン化はタンパク質修飾の一種で、ユビキチンリガーゼなどの酵素の働きによりユビキチンがイソペプチド結合で基質タンパク質に付加されることを指す。
(注8)翻訳伸長
リボソームがmRNAの読み枠内のコドンに従ってアミノ酸をつなげていく過程。翻訳伸長は通常開始コドンで始まり、終止コドンで完結する。
(注9)質量分析
物質を微細なイオンにし、その質量電荷比によって分離・検出することで物質の同定や定量を行う手法である。
")
