2023-08-30 森林総合研究所,新潟大学,東京大学,基礎生物学研究所,新潟県
ポイント
- 花粉症対策に光明!スギ花粉の生産に関わる遺伝子を新たに特定
- この遺伝子のたった1塩基の変異で花粉ゼロ
- 無花粉スギの苗木生産の増大につながる成果
概要
国立研究開発法人森林研究・整備機構森林総合研究所、新潟大学、東京大学大学院新領域創成科学研究科、基礎生物学研究所、新潟県森林研究所の研究グループは、無花粉スギの原因遺伝子MS4を特定しました。人工交配によって作製したスギ集団を用いた遺伝分析とスギ参照ゲノム配列を活用して候補となる遺伝子を絞り込みました。得られた候補遺伝子が花粉生産に関わることをモデル植物のシロイヌナズナを用いて検証することに成功しました。MS4は、花粉壁の生成に関わる酵素(TKPR1)を合成する遺伝子であり、この遺伝子のわずか1つの塩基が変異することで無花粉になることが明らかになりました。現在、無花粉スギは、MS4とは別の遺伝子が変異した無花粉スギの雌花に有花粉のスギをかけあわせて無花粉スギの種子を得て苗木を生産していますが、この方法においてMS4を使用する選択肢が得られたことで無花粉スギの新たな育種や生産の増大につながると想定され、花粉症対策への貢献が見込まれます。
本研究成果は、2023年8月8日にPNAS Nexus誌でオンライン公開されました。
背景
スギ花粉症は、我が国の深刻な社会問題となっています。スギ花粉症患者は年々増加し、日本人の約4割が罹患しているとする調査結果もあります。林業分野におけるスギ花粉症対策の1つは花粉発生源を減少させることです。政府は、花粉発生源対策として、スギ人工林を減少させるとともに、無花粉スギ(用語解説1)を含む花粉の少ないスギ苗木への植え替えを促進するなどして、30年後に花粉発生量を半減させる目標を掲げています。特に無花粉スギへの植え替えは、森林のCO2吸収機能を維持しつつ花粉を飛散させずに木材資源を確保できる非常に有効な策であると言えます。近年無花粉スギの原因遺伝子のひとつであるMS1(用語解説2)が特定され、無花粉スギと有花粉のスギをかけあわせて得られる無花粉スギの種子を使った苗木の生産(用語解説3)に活用されています。無花粉スギの遺伝子はMS1以外にも3つの遺伝子(MS2、MS3および MS4)の存在が知られており、MS1やこれらの遺伝子を活用することで、無花粉スギの苗木の生産効率をさらに高めることができますが、これまでMS1以外の遺伝子の正体は不明でした。
内容
今回研究グループは、花粉が成熟する直前に異常が生じて無花粉になるタイプ(MS4)に着目しました。このタイプの無花粉スギの雄花断面を観察すると、ごく少量の花粉しか確認できず、電子顕微鏡で詳細に観察すると、異常な形の花粉が観察されるとともに、花粉外壁の外層構造が不完全であることがわかりました(図1)。すなわちMS4タイプの無花粉スギは、花粉を作らないMS1タイプとは異なり、機能が不全かつ飛散しにくい花粉を作ることがわかりました。
次に、MS4の原因遺伝子を特定するために、人工交配による遺伝分析と2023年に森林総合研究所を中心とするグループが解読を完了して公開したスギの参照ゲノム配列(用語解説4)を利用して、MS4遺伝子が存在する領域を第4染色体の765万塩基対の範囲に絞り込みました。この範囲に、スギの雄花で活発に働く遺伝子が1つだけ発見されました。その遺伝子の塩基配列を解読したところ、花粉壁の形成に関わる酵素(TKP1、用語解説5)を合成すると予想されました。興味深いことに無花粉スギのTKPR1遺伝子は、DNAたった1塩基の置換に由来するアミノ酸変異(82番目のアミノ酸のシステイン(C)がアルギニン(R)に置換する変異であることからC82Rと呼ばれます)を持つことがわかり、この変異がTKPR1遺伝子の機能に影響する可能性が考えられました(図2)。
TKPR1遺伝子は多くの植物で働いていることが知られています。そこで、この1塩基の変異が花粉生産の有無を決めるかどうかを確かめるためにモデル植物のシロイヌナズナ(用語解説6)の突然変異体を用いて実験しました。TKPR1が欠損した無花粉のシロイヌナズナに正常なスギのTKPR1遺伝子を導入すると花粉が生産された一方で、C82Rの変異を持つTKPR1遺伝子を導入しても正常な花粉は生産されませんでした(図3)。このことからTKPR1がわずか1つの塩基置換により機能を喪失することが初めて証明されました。そして、MS4の原因遺伝子はTKPR1遺伝子であることが証明されました。

図1 正常なスギ(左側)とMS4タイプの無花粉スギ(右側)の雄花および花粉の形態比較
スギの雄花の房(左上)から雄花を1つ取りナイフで切断しました。正常なスギでは非常に多く粉状の花粉が確認できますが(A)、無花粉スギではそれを確認できません(B)。花粉を電子顕微鏡で観察すると、正常なスギは楕円体にパピラと呼ばれる突起(赤い矢印の部分)のついた特徴的な形の花粉の粒子が観察されますが(C)、無花粉スギでは粒子がつぶれており、飛散する能力がないと考えられます(D)。また正常な花粉の表面にはオービクルと呼ばれる粒子状の構造(白い矢印)が全体的に多く観察できますが(C)、無花粉スギでは数が少なく形も異常でした(D)。さらに花粉の切断面を電子顕微鏡で観察してみると正常なスギでは花粉外壁の外層(Ec)が十分に発達している(E)一方で、無花粉スギでは未発達(dEc)であることがわかりました(F)。なお花粉症の原因となる物質(「抗原」と呼びます)は花粉外壁の外層(Ec)やオービクル等に存在することが知られています。
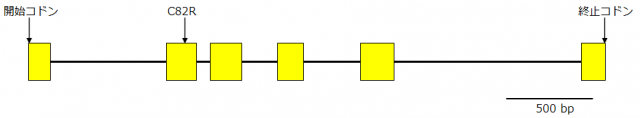
図2 MS4の遺伝子構造
MS4の構造を表す模式図です。長方形の部分がタンパク質を合成する部分を、長方形と長方形を結ぶ線分がイントロン(遺伝子内に存在し、タンパク質を合成せず、RNA合成時に取り除かれる領域)と呼ばれる部分を表します。MS4は5つのイントロンで分断されています。図では左端に開始コドン、右端に終止コドンを示し、3,323塩基対(bp)、333アミノ酸残基に対応する部分を示しています。MS4が合成するタンパク質は花粉外壁の主要な成分である脂質の生合成に必要な酵素(TKPR1)と予想されました。無花粉スギではTKPR1の82番目のアミノ酸のシステイン(C)がアルギニン(R)に置換する変異(C82R)が存在し、この変異は酵素の機能に影響する有害な変異であると予想されました。
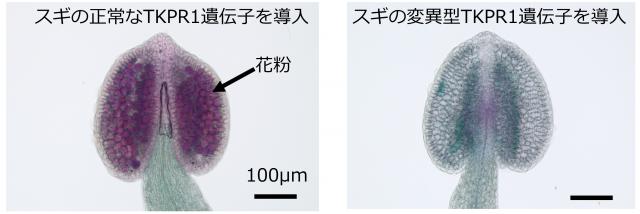
図3 MS4の遺伝子機能証明実験におけるシロイヌナズナ雄しべの顕微鏡写真
シロイヌナズナのTKPR1機能欠損変異体(TKPR1が機能せず無花粉の変異体)に正常なスギのTKPR1遺伝子を導入すると花粉が生産され稔性が回復します(左図)。図では染色液により花粉は赤紫色に染まっています(矢印)。一方で、同じシロイヌナズナ変異体に、C82Rの変異を持つ変異型TKPR1遺伝子(正常なスギのTKPR1遺伝子とは1塩基だけ異なる塩基配列)を導入しても無花粉のままでした(右図)。このことにより無花粉スギのMS4の正体はTKPR1遺伝子であり、この遺伝子のわずか1塩基の違いが花粉生産の有無を決定することが明らかになりました。
今後の展開
本研究により、MS4タイプの無花粉スギの原因遺伝子がTKPR1であること、そしてTKPR1遺伝子のわずか1塩基の違いが花粉の生産を決定することが明らかになりました。現在無花粉スギの苗木を生産するにはMS1と呼ばれる遺伝子が実際に活用されています。MS1をホモ接合で保有するスギとヘテロ接合で保有するスギをかけあわせて得られる種子から育てた苗木を無花粉スギとして出荷しています。無花粉スギの育種や生産に際してMS1に加えてMS4も利用することで、苗木の生産の増大が見込まれます。さらにTKPR1遺伝子は花粉を作る植物が共通して有する遺伝子であるため、今後TKPR1遺伝子を利用することでスギ以外のさまざまな植物で花粉症対策を行えることも期待されます。
用語解説
(1)無花粉スギ
花粉の形成に異常があり正常な花粉ができないため花粉を飛散せず、林業用に植栽するスギを林業用語で無花粉スギと呼んでいます。科学用語では雄性不稔スギと呼びますが、雄性不稔には様々なタイプがあり、今回のMS4タイプは花粉の粒子が観察できる雄性不稔です。花粉が飛散しないため花粉症対策として活用できます。
(2)MS1
無花粉スギの原因遺伝子の1つで、最初に見つかった遺伝子であるため MS1と命名されています。雄性不稔を意味する英語(male sterility)の頭文字に由来します。MS1は脂質を輸送する機能があると推定されています(http://www.ffpri.affrc.go.jp/pubs/seikasenshu/2021/documents/p38-39.pdf)。
(3)無花粉スギの種子を使った苗木の生産
現在、無花粉スギの種子は付図1で示すような人工交配により生産されています。この図では正常な(野生型の)MS1遺伝子を「A」、変異型のMS1遺伝子を「a」で表しています。無花粉スギは変異型の遺伝子を2つ持っています(aaのホモ接合と呼びます)。無花粉スギでも雌花は正常に咲くので種子親になれます。花粉親には野生型遺伝子Aと変異型遺伝子aを1つずつ持っている(Aaのヘテロ接合と呼びます)スギを選びます。変異型遺伝子aを持っていてもヘテロ接合の場合は花粉ができるので花粉親になれます。そしてこのAaの花粉親とaaの無花粉スギの種子親との交配によりできた種子には、無花粉スギ(aa)と有花粉スギ(Aa)がほぼ等量ずつ含まれ、約50%が無花粉スギ(aa)の苗木となります。
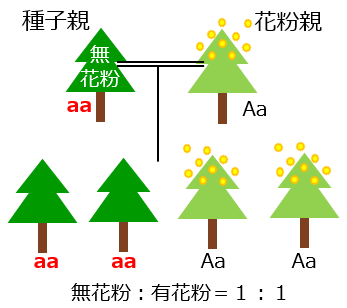
付図1 無花粉スギの種子による苗木の生産
(4)参照ゲノム配列
種を代表するゲノム配列で、この配列を基準にして近縁種間で配列を比較したり、種内の品種間の配列を比較したりするなどの高度な解析を統一的に行うことができます。スギの参照ゲノム配列は近年の分析技術の進歩により解読され(https://www.ffpri.affrc.go.jp/press/2023/20230301/index.html)、今回のMS4遺伝子の同定で活用されました。
(5)TKPR1
テトラケチドα-ピロンリダクターゼ1(Tetraketide alpha-pyrone reductase 1)は葯の内側にあるタペートと呼ばれる組織で働く酵素で、花粉外壁の主要な成分である脂質の生合成に関与します。合成された脂質は花粉外壁の外層に蓄積します。本文ではローマン体で統一していますが、論文中では遺伝子名(TKPRI1:イタリック体)とその産物の酵素タンパク質(TKPR1:ローマン体)を区別して書いています。
(6)シロイヌナズナ
小型の草本植物でモデル生物として科学研究で広く使われています。世代期間が短いために遺伝学的な研究もしやすく、遺伝子を人為的に操作する「形質転換」などの技術を容易に行うことができます。
論文
タイトル:A single-nucleotide substitution of CjTKPR1 determines pollen production in the gymnosperm plant Cryptomeria japonica
著者:Hiroyuki Kakui, Tokuko Ujino-Ihara, Yoichi Hasegawa, Eriko Tsurisaki, Norihiro Futamura, Junji Iwai, Yuumi Higuchi, Takeshi Fujino, Yutaka Suzuki, Masahiro Kasahara, Katsushi Yamaguchi, Shuji Shigenobu, Masahiro Otani, Masaru Nakano, Masaaki Nameta, Shinsuke Shibata, Saneyoshi Ueno, Yoshinari Moriguchi
掲載誌:PNAS Nexus
論文URL:https://doi.org/10.1093/pnasnexus/pgad236(外部サイトへリンク)
研究費:森林総合研究所交付金プロジェクト「有用遺伝子の特定に向けたスギ全ゲノム走査(201421)」、「スギの基盤遺伝情報の高精度化と有用遺伝子の機能解明(201906)」、農林水産省 農林水産業・食品産業科学技術研究推進事業および生研支援センター イノベーション創出強化研究推進事業(JPJ007097)「無花粉スギの普及拡大に向けたDNAマーカー育種技術と効率的な苗木生産技術の開発(Project ID 28013B)」、文部科学省科学研究費補助金「花粉数を制御する遺伝子ネットワークの解明と作物への応用(19K05976)、植物の巨大なゲノムを解読・解析する手法(20H03239)」、「植物の巨大なゲノムを解読・解析する手法(20H03239)」、「林木初のピラミッディング育種技術の高度化と実用化に向けた検証(21K05666)」、「稔性が回復する雄性不稔遺伝子の同定と生理生態(23H02263)」、基礎生物学研究所共同利用研究「スギの全ゲノム配列の解読(16-403, 17-405, 18-408)」
共同研究機関
森林総合研究所、新潟大学、東京大学、基礎生物学研究所、新潟県森林研究所
報道機関関係者の方々へのお願い
本研究に興味を持っていただきありがとうございます。本研究成果を取り上げる際には、読者や聞き手が研究の詳細を知り、結果の背景を正確に理解することのできるよう原典の論文を引用していただきますようお願い致します。特にウェブサイト版での記事やSNS(TwitterやFacebook、YouTube等)等での情報発信の際には、こちらの論文URL(https://doi.org/10.1093/pnasnexus/pgad236)を付けていただくことを検討いただければ幸いです。また、このお願いにつきまして生物科学学会連合から提出されました「研究成果をメディアへ報道する際のお願い」(https://seikaren.org/wp/wp-content/uploads/2023/02/to-media.pdf (外部サイトへリンク)も併せてご覧いただければ幸いです。
また英文によるプレスリリースも掲載誌(PNAS Nexus)から行われていることを申し添えます(https://www.eurekalert.org/news-releases/997816(外部サイトへリンク))。
お問い合わせ先
研究担当者:
森林総合研究所 樹木分子遺伝研究領域 樹木遺伝研究室 チーム長 上野真義
広報担当者:
森林総合研究所 企画部広報普及科広報係
")
