2023-01-20 理化学研究所
理化学研究所(理研)環境資源科学研究センター 合成ゲノミクス研究グループの栗原 志夫 研究員(研究当時、現 客員研究員)、赤木 千佳 研修生、松井 南 グループディレクターらの共同研究チームは、植物では、翻訳装置であるリボソーム[1]の生合成などに関わる遺伝子群の翻訳の活性化に「青色光を情報として正常に伝達する機能」と「正常な葉緑体機能」の両方が必要であることを明らかにしました。
本研究成果は、植物の新たな成長制御の解明につながると期待できます。
これまで、青色光を受容することによる植物の遺伝子発現は転写と翻訳の両レベルで制御されていることが分かっていましたが、翻訳レベルの発現変動がどのように制御されているかは分かっていませんでした。
今回、共同研究チームは、通常の培地にまいた青色光を受容できなくさせたシロイヌナズナの変異体の種子と、葉緑体機能阻害剤を添加した培地にまいた野生型の種子をそれぞれ暗所にて生育させた後、青色光で露光したときの遺伝子の発現変化をゲノム全域にわたって調べました。その結果、変異体の芽生えと葉緑体機能阻害剤下で生育した芽生えとでは、それぞれのコントロールでは発現していたリボソームの生合成などに関わる遺伝子が発現しないことを発見しました。これにより、植物における翻訳レベルでの遺伝子発現制御には、青色光を正常に伝達する機能と正常な葉緑体機能の二つが必要であることが分かりました。このことは、植物のリボソームの生合成などに関わる遺伝子の翻訳の活性化には光を受容することが必要であることを示しています。
本研究は、科学雑誌『Journal of Plant Research』オンライン版(1月20日付)に掲載されました。
背景
植物の種子は土壌中で発芽した後、胚軸を地上へと伸ばします。地上に出た芽生えは、光を受容することにより子葉を展開します。子葉内では、光合成を行う葉緑体が発達していきます。この初期の光受容は遺伝子発現に劇的な変化を起こしますが、その制御機構の詳細については未知な部分が多く残っています。
遺伝子発現とは、ゲノム上の遺伝子の情報(塩基配列)がメッセンジャーRNA(mRNA)[2]に転写され、mRNAが翻訳装置であるリボソームに運ばれて、タンパク質に翻訳されることです。近年、網羅的な転写と翻訳の解析が可能になったことから、それぞれの遺伝子がコードするmRNAの蓄積量とタンパク質の翻訳量は必ずしも比例せず、タンパク質の翻訳量はmRNAの蓄積量に依存することなく増減していることが明らかになりました。
2020年に松井グループディレクターらは、暗所にて発芽したシロイヌナズナの芽生えに、植物の成長に関わる主要な光波長である青色光を照射したときの核遺伝子の転写産物量と翻訳量の初期変化について報告しました注1)。このとき、青色光を受容することによる遺伝子発現の制御が転写レベルだけではなく、翻訳レベルでも行われていることが明らかになりました。しかし、その翻訳レベルの発現変動がどのように制御されているかは分かっていませんでした。
注1)2020年3月16日プレスリリース「光受容によるタンパク質の翻訳変化を解明」
研究手法と成果
共同研究チームはまず、青色光を受容できないシロイヌナズナのcry1cry2二重変異体[3]およびhy5変異体[4]の種子を通常の培地にまいた場合と、野生型(WT)の種子を葉緑体機能阻害剤(DCMUおよびNorflurazon)が添加された培地にまいた場合で、それぞれを暗所で生育させた後に青色光を露光したときの、芽生えの前後での翻訳量の変化を、ゲノム全域にわたってリボソームプロファイリング解析(Ribo-seq解析)[5]で調べました。
翻訳レベルで発現制御されている遺伝子群を明らかにするために、mRNA 1分子当たりのリボソーム量を翻訳効率として算出しました。翻訳効率が上昇していた遺伝子がどのような機能を持つのかを調べるために、それらのリストを使用して遺伝子オントロジーエンリッチメント解析(GOエンリッチメント解析)[6]を行いました。その結果、青色光を受容できなくさせた変異体のコントロールとなるWTの芽生えや葉緑体機能阻害剤のコントロールとなるDMSO添加条件下における芽生えの遺伝子リストには、リボソームの生合成などに関わる遺伝子が多く含まれていましたが、変異体および葉緑体機能阻害剤下で生育させた芽生えの遺伝子リストにはリボソームの生合成などに関わる遺伝子が含まれていないことが分かりました。
そこで、コントロールで認められたリボソームの生合成などに関わる遺伝子の翻訳効率が、変異体および葉緑体機能阻害剤下で生育させた芽生えとコントロールとで差があるのかをヒートマップ[7]を作成して確認しました。すると、変異体および葉緑体機能阻害剤下で生育させた芽生えは、コントロールに比べて翻訳効率が低下していることが分かりました(図1)。この結果は、GOエンリッチメント解析において、リボソームの生合成などに関わる遺伝子が変異体および葉緑体機能阻害剤下で生育させた芽生えの遺伝子リストに認められなかったことと一致しています。

図1 リボソーム生合成に関わる遺伝子の翻訳効率の比較
翻訳効率をヒートマップで表した。コントロールと比較して、変異体の芽生え(A)と葉緑体機能の阻害剤を添加した培地にて生育させた芽生え(B)では、翻訳効率が低下していた。
これらの結果から、植物が光を初めて受容したときにリボソームの生合成などに関わる遺伝子の翻訳を活性化させるには、「青色光を情報として正常に伝達する機能」と「正常な葉緑体機能」の両方が必要であることが明らかになりました(図2)。このことは、光に反応することで、リボソームの構成タンパク質が翻訳レベルで増えていることを示しています。植物は光を受容して形態形成を進めるため、形態形成に必要な多くの遺伝子の発現を飛躍的に上昇させる必要があります。従って、リボソームの生合成などに関わる遺伝子の発現を翻訳レベルで上昇させることは、植物にとって合理的な戦略であると考えられます。
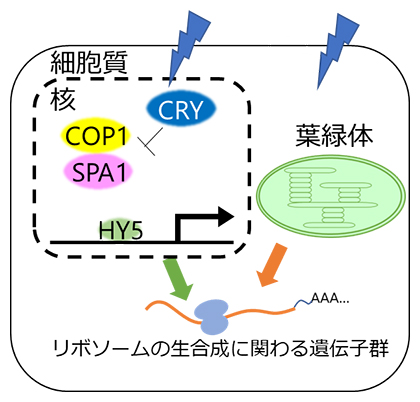
図2 青色光を受容したときのリボソーム生合成関連遺伝子群の翻訳を活性化させるシグナル
受容体CRY(クリプトクロム)と転写因子HY5が存在する場合(WT)は、青色光照射によってCRYが活性化される。その影響でタンパク質分解に関わるCOP1/SPA1タンパク質複合体の活性が低減し、HY5は分解されずに核コードの光応答性遺伝子の転写を誘導する。このように、青色光シグナルを正常に伝達する機能がリボソームの生合成などに関わる遺伝子群の翻訳の活性化には必要であり、また、正常な葉緑体機能も必要である。
今後の期待
本研究では、シロイヌナズナの芽生えが初めて光を受容したときの遺伝子の発現変化を調べることで、リボソームの生合成などに関わる遺伝子の翻訳の活性化には、青色光を情報として正常に伝達する機能と正常な葉緑体の機能との両方が必要であることを明らかにしました。
これまでは、シロイヌナズナの芽生えが青色光を初めて受容したときの翻訳制御に何が関わっているのか、というところまでは研究されていませんでしたが、今回の研究結果は、植物のリボソームの生合成には光を受容することも必要であることを示しています。従って、本研究成果は植物の新たな成長制御の解明につながると期待できます。
また本研究成果は、国際連合が2016年に定めた17項目の「持続可能な開発目標(SDGs)[8]」のうち、「7.エネルギーをみんなにそしてクリーンに」、「13.気候変動に具体的な対策を」、「15.陸の豊かさも守ろう」に貢献するものです。
補足説明
1.リボソーム
細胞の小器官の一つ。メッセンジャーRNA(mRNA)から情報を読み取り、タンパク質へ翻訳する。転写されたmRNAはリボソームに結合する。そこにtRNA(トランスファーRNA)が遺伝子情報に対応したアミノ酸を運び、タンパク質が作られる。
2.メッセンジャーRNA(mRNA)
タンパク質は核内のDNA配列の遺伝子にあたる部分がRNAという形でコピーのように写し取られ(転写)、それが読み出される(翻訳)ことで作られる。この転写されたRNAをメッセンジャーRNAと呼ぶ。タンパク質の翻訳はリボソームで行われる。
3.cry1cry2二重変異体
植物は青色光と紫外線を受容するときに、クリプトクロム(CRY)を受容体として使用している。シロイヌナズナには、CRY1とCRY2が主に青色光受容体として機能している。その機能を欠損させた変異体がcry1cry2二重変異体である。
4.hy5変異体
光を情報として遺伝子に伝える転写因子として、HY5(ELONGATED HYPOCOTYL 5)がある。HY5転写因子はクリプトクロムなどの受容体から得た光を情報として受け取り、HY5転写因子よりも下流で働く光応答性の遺伝子の転写を促す。その転写因子の機能を欠損させた変異体がhy5変異体である。
5.リボソームプロファイリング解析(Ribo-seq解析)
組織からリボソームを抽出し、リボソームと結合しているRNA配列(リボソームフットプリント)を同定することにより、どの遺伝子がどの程度の効率で翻訳されているのかを調べる解析手法。
6.遺伝子オントロジーエンリッチメント解析(GOエンリッチメント解析)
遺伝子に付与される注釈のことを遺伝子オントロジー(gene ontology:GO)といい、遺伝子の生物的プロセス、細胞の構成要素、分子機能に分類される。遺伝子オントロジーエンリッチメント解析は、ある遺伝子セットの中に、特定の遺伝子オントロジーを持つ遺伝子が多く含まれているか否かを調べる手法。
7.ヒートマップ
遺伝子発現のデータを視覚化する手法。遺伝子発現の高低を色で示す。今回は、遺伝子発現の高いところを緑色、低いところをピンク色で示している。
8.持続可能な開発目標(SDGs)
持続可能な開発目標(SDGs)とは、2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標である。持続可能な世界を実現するための17のゴールから構成され、地球上の誰ひとりとして取り残さないことを誓っている。SDGsは発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に取り組んでいる。(外務省のホームページから一部改変して転載)
共同研究チーム
理化学研究所 環境資源科学研究センター 合成ゲノミクス研究グループ
グループディレクター 松井 南(マツイ・ミナミ)
研究員(研究当時、現 客員研究員)栗原 志夫(クリハラ・ユキオ)
(東京大学大学院 総合文化研究科 助教)
研究員(研究当時、現 客員研究員)蒔田 由布子(マキタ・ユウコ)
(前 橋工科大学 工学部 生命情報学科 教授)
研修生 赤木 千佳(アカギ・チカ)
京都大学大学院 理学研究科 生物科学専攻 生体分子情報領域
教授 青山 卓史(アオヤマ・タカシ)
准教授 柘植 知彦(ツゲ・トモヒコ)
研究支援
本研究は、RIKEN Pioneering Projects「Biology of Intracellular Environments(研究分担者:松井南)」の支援を受けて実施し、日本学術振興会(JSPS)科学研究費助成事業基盤研究(B)「植物の成長段階における転写・翻訳開始点に依存したmRNAの運命決定の解明(研究代表者:栗原志夫)」による助成を受けて行われました。
原論文情報
Chika Akagi, Yukio Kurihara, Yuko Makita, Masaharu Kawauchi, Tomohiko Tsuge, Takashi Aoyama, Minami Matsui, “Translational activation of ribosome-related genes at initial photoreception is dependent on signals derived from both the nucleus and the chloroplasts in Arabidopsis thaliana.”, Journal of Plant Research, 10.1007/s10265-022-01430-8
発表者
理化学研究所
環境資源科学研究センター 合成ゲノミクス研究グループ
グループディレクター 松井 南(マツイ・ミナミ)
研究員(研究当時、現客員研究員)栗原 志夫(クリハラ・ユキオ)
研修生 赤木 千佳(アカギ・チカ)
報道担当
理化学研究所 広報室 報道担当
〜")
