2023-10-20 東京大学
森 秀世(生物科学専攻 博士課程)
稲垣 宗一(生物科学専攻 准教授)
角谷 徹仁(生物科学専攻 教授)
発表のポイント
- ヒストン脱メチル化酵素のLDL3タンパク質が転写複合体と協働し、転写されている遺伝子からヒストン修飾のH3K4me2を除去していることを見出しました。
- LDL3は植物の発生や再生に重要であり、そこにヒストン脱メチル化を介した「転写の記録」が役割を果たすと考えられます。
- 多くの遺伝子の発現制御に関わるH3K4メチル化の制御メカニズムについて理解を深め、環境変化に強い植物の作出などへの応用も期待できます。

LDL3はRNAポリメラーゼIIと結合し転写共役的にH3K4me2を除く
発表概要
東京大学大学院理学系研究科の森秀世大学院生、稲垣宗一准教授、角谷徹仁教授らのグループは、モデル植物であるシロイヌナズナ(注1)において、H3K4me2(注2)特異的な脱メチル化酵素であるLDL3が転写装置(注3)と協働することで、転写活性遺伝子からH3K4me2を除去することを明らかにしました。陸上植物においてH3K4me2の蓄積量は遺伝子発現レベルと負の相関を示すことが報告されており、その負の相関を形成する分子機構を示す結果です。植物は、転写した遺伝子をH3K4me2が少ない状態としてゲノム上に記録し、遺伝子発現を頑健に制御していると考えられます。
先行研究からLDL3は植物の発生や再生といった発生可塑性(注4)の発揮に重要であることが知られており、本研究は植物の発生可塑性の根底にある分子機構の一つを明らかにするものです。環境変化に強い植物の作出などへの応用も期待できます。
発表内容
H3K4meは真核生物に広く保存されたクロマチン修飾(注5)であり、ゲノムの中でも特に発現レベルの高い遺伝子のコード領域にみられることから、転写活性化修飾として一般に考えられています。確かに、H3K4me3レベルは酵母、動物、植物に共通して、遺伝子発現レベルと正の相関を示しますが、H3K4me2レベルは陸上植物においては遺伝子発現レベルと負の相関を示し、転写抑制的修飾であることを示唆する結果が報告されていました。しかし、陸上植物において遺伝子発現レベルと負の相関を示すようなH3K4me2のゲノム内分布パターンがどのように形成されるのか、その分子機構は未解明でした。
研究チームはまず、モデル植物のシロイヌナズナにおいて、H3K4me2を特異的に除去する脱メチル化酵素であるLDL3に注目しました。LDL3はゲノム上のH3K4me2を除去しているため、その機能欠損変異体ではH3K4me2が野生型に比べ多く蓄積します。しかし、その影響は全ての遺伝子に均等ではなく、LDL3がH3K4me2を除去するターゲット遺伝子とそれ以外の遺伝子があることを見出しました。LDL3ターゲット遺伝子がどのように決定されているのかを探るため、機械学習によってLDL3ターゲット遺伝子の特徴を探索したところ、LDL3ターゲット遺伝子はリン酸化されたRNAポリメラーゼIIが多く局在する遺伝子であることがわかりました。遺伝子の転写装置であるRNAポリメラーゼIIのリン酸化は転写活性と結びついており、LDL3は転写が活発な遺伝子領域でH3K4me2を除去していると考えられます。
次に、植物体からLDL3タンパク質複合体を回収し、LDL3と結合しているタンパク質を調べました。その結果、LDL3はRNAポリメラーゼIIと結合しており、さらに、リン酸化を受けたRNAポリメラーゼIIとより強く結合していることがわかりました。
また、転写伸長因子(注6)の機能欠損変異体を用いた遺伝学的な解析の結果、転写伸長に異常が生じると、LDL3のゲノム上の局在が変化し、その結果H3K4me2パターンも乱れてしまうことを見出しました。これらの結果は、転写伸長中のRNAポリメラーゼII複合体がそのリン酸化を介してLDL3を呼び込み、転写領域のH3K4脱メチル化を促していることを示しています(図1)。
図1:LDL3はRNAポリメラーゼIIと結合し転写共役的にH3K4me2を除く
最後に、研究チームはldl3機能欠損変異体において、H3K4me2レベルと遺伝子発現レベルの負の相関が崩れ、正の相関に近づくことを見出しました(図2)。この結果は、植物にユニークなH3K4me2レベルと遺伝子発現レベルの負の相関がLDL3によって形成されていることを示唆します。LDL3が転写装置と協働してH3K4me2を除く結果、転写が活発な遺伝子にはH3K4me2が少なく、あまり転写されていない遺伝子にH3K4me2が残る分布パターンが出来上がるのだと考えられます。
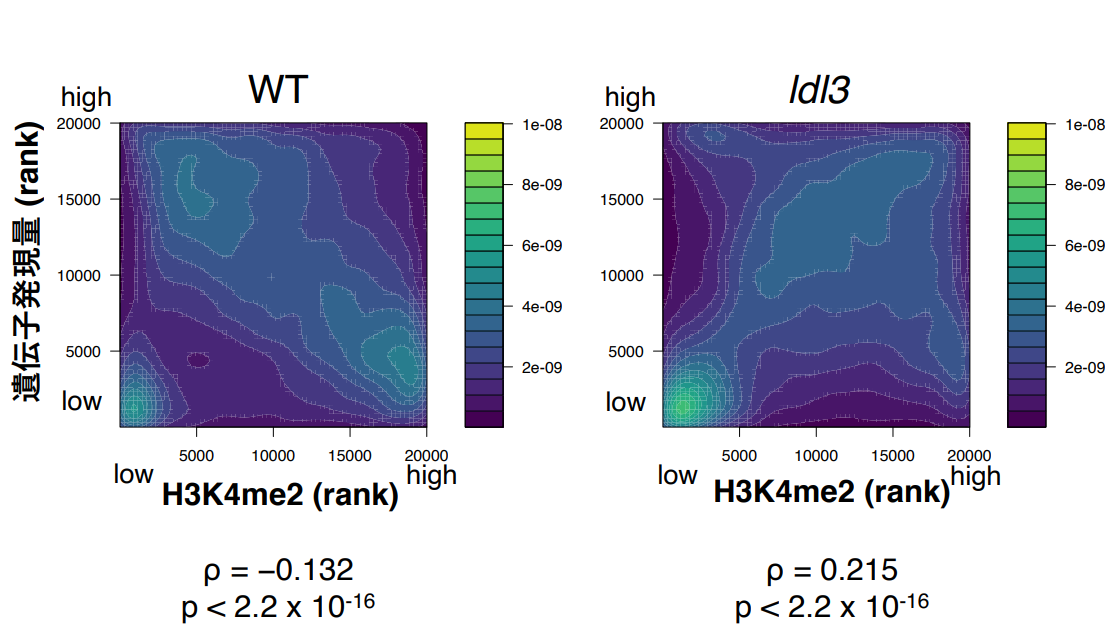
図2:H3K4me2と遺伝子発現量との相関
横軸にH3K4me2レベル、縦軸に遺伝子発現量を示し、遺伝子密度をヒートマップで表した。
左 : 野生型、右 : ldl3変異体。ρは順位相関係数。
植物は環境に応じて可塑的に発生を制御することができます。環境に応じて遺伝子発現状態を変化させる際、LDL3によって形成されるH3K4me2の分布パターンは、転写状態の記録として機能し、新たな転写状態を確立するのに役立つと考えられます。実際、先行研究では、LDL3が開花時期や植物体の再生の制御に重要であることが示されています。本研究は、植物の発生可塑性制御の根底にある分子機構の一つを明らかにしました。LDL3が担う発生可塑性は植物の重要な環境適応能力であり、農学や環境問題対応など応用科学への波及効果も期待できます。
論文情報
- 雑誌名
EMBO Journal論文タイトル
Cotranscriptional demethylation induces global loss of H3K4me2 from active genes in Arabidopsis著者
Shusei Mori*, Satoyo Oya, Mayumi Takahashi, Kazuya Takashima, Soichi Inagaki* , Tetsuji Kakutani*DOI番号
10.15252/embj.2023113798
研究助成
本研究は、科研費(課題番号20H05913,21H04977,22H02299,23H00365)、HFSP (RGP0025/2021)、さきがけ (JPMJPR17Q1) 、CREST (JPMJCR15O1)の支援により実施されました。
用語解説
注1 シロイヌナズナ
アブラナ科の一年草で、世代時間が短い、ゲノムサイズが小さいなどの利点から植物遺伝学の代表的なモデル生物となった。真核生物に存在するクロマチン修飾のほとんどを持つことから、クロマチン修飾の研究も活発に行われている。
注2 H3K4me2
ヒストンH3タンパク質の4番目のリジンのメチル化をH3K4meと示し、メチル基の数によりme1, me2, me3と表記した。H3K4me2はリジンに2つメチル基がついた状態のこと。
注3 転写装置
RNAポリメラーゼを含むタンパク質複合体で、DNAを鋳型にRNAを合成する(転写する)装置。
注4 発生可塑性
生物が生育環境に応じて、柔軟に発生プログラムを変える性質。例えば植物は、気温や日長を感知して発生様式を変化させることで、適切な時期に開花する。
注5 クロマチン修飾
真核生物のDNAはヒストンなどと結合してヌクレオソームを形成し、さらにヌクレオソームが折りたたまれてクロマチン構造を形成している。このクロマチンに付加する化学修飾をまとめてクロマチン修飾と呼び、DNAのメチル化やヒストンのメチル化、アセチル化などが知られている。
注6 転写伸長因子
RNAポリメラーゼIIと結合し、転写伸長を制御しているタンパク質。

