
2019-08-28 埼玉大学, 基礎生物学研究所
【概要】
埼玉大学大学院理工学研究科 生命科学系専攻 生体制御学コースの大学院生 伴 博之(平成30年度 博士前期課程修了)、横田大佑(平成27年度 博士前期課程修了)と川村哲規 准教授を中心とする研究グループは、埼玉大学大学院理工学研究科 弥益 恭 教授、自然科学研究機構 基礎生物学研究所 高田慎治 教授、矢部泰二郎 助教と共同で、脊椎動物のからだに見られる繰り返し構造のもととなる体節が発生過程で迅速に形成される分子メカニズムの一端を、小型熱帯魚ゼブラフィッシュを用いて明らかにしました。この研究成果は、発生生物学分野における重要な研究成果が掲載される英国国際学術専門誌「Development」に8月23日付のオンライン版で掲載されました。
【研究内容】
ヒトを始めとする脊椎動物のからだには、類似した構造が反復した繰り返し構造がみられます。例えば、脊椎動物が共通にもつ背骨は脊椎骨と呼ばれる骨が重なった繰り返し構造をしています。他にも、筋肉や神経、血管にも、このような繰り返し構造がみられます。この繰り返し構造はもとを正すと、からだが形作られる発生の過程で生じる「体節」と呼ばれる組織に由来します。体節が数十個、連なって正しく形成された結果、成体の繰り返し構造が出来上がるのです(図1)。発生の過程で、様々な組織や器官が形成されますが、体節は他と比べ、迅速に形成されます。例えば、今回の研究で用いたゼブラフィッシュでは約30分間隔で形成されますが、体節がどのようにして、このように迅速に形成されるのかについては不明でした。
 図1 発生過程で生じた体節の繰り返し構造が、成体のからだの繰り返し構造のもととなる。体節は、脊椎動物の胚発生時に共通にみられる構造。
図1 発生過程で生じた体節の繰り返し構造が、成体のからだの繰り返し構造のもととなる。体節は、脊椎動物の胚発生時に共通にみられる構造。
脊椎動物のからだに分節構造をもたらす体節は、発生過程において胚の後方に位置する未分節中胚葉の前方が周期的にくびれ切れることで形成されます。未分節中胚葉において次の予定体節境界の位置(分節ポイント)が正しく決定されることが、正常な体節形成に必須です。マウス胚やゼブラフィッシュ胚を用いた解析から、分節ポイントを決定するメカニズムとして、未分節中胚葉で特異的に発現するT-box型転写因子 Tbx6タンパク質の前方発現境界が次の分節ポイントを決定する最も重要な要素であることが示されていました(図1)。実際に、ゼブラフィッシュtbx6変異体では、体節境界が形成されません。このように、Tbx6タンパク質の前方境界の制御は、体節形成において分節ポイントを規定する普遍的な機構であると考えられます。またTbx6タンパク質は、川村准教授らが同定したRipply (Kawamura et al., Developmental Cell, 2005)により分解されることで、その前方境界が後方へシフトし、次の体節の境界になることが示されていました(図2)。このように、体節が迅速に、正しく形成されるメカニズムを理解するためには、このTbx6タンパク質の前方境界がどのようにして効率的に後方へシフトするのかを理解することが必要です。そのためにはRipplyによりTbx6タンパク質が分解される機構とともに、Tbx6タンパク質を生じる源であるtbx6遺伝子の転写がどのようにして適切に抑制されるかを理解する必要があります。
 図2 Tbx6タンパク質の前方境界が、次の分節ポイントとなる。Tbx6タンパク質は、Ripplyにより分解され、体節一つ分、後方へシフトする。これが繰り返され、前後に連なった体節の繰り返し構造が出来上がる。
図2 Tbx6タンパク質の前方境界が、次の分節ポイントとなる。Tbx6タンパク質は、Ripplyにより分解され、体節一つ分、後方へシフトする。これが繰り返され、前後に連なった体節の繰り返し構造が出来上がる。
本研究ではまず、tbx6遺伝子の転写制御の重要性に着目し、ゼブラフィッシュを用いてtbx6遺伝子の転写制御に関わるゲノム領域を探索し、発現制御領域TR1を同定しました。TR1領域を詳しく調べた結果、Tbx6タンパク質が結合する配列が存在し、実際にTbx6タンパク質がTR1領域に結合することが分かりました。ゲノム編集技術によりその制御領域を欠失した変異体を単離したところ、tbx6制御領域を欠いた変異体では、ゼブラフィッシュ約30対の体節のうち、7~25番目の体節境界が形成されませんでした(図3)。この表現形は体節境界が全く形成されないtbx6変異体と類似しており、さらにtbx6制御領域変異体において、tbx6 mRNAが発生時期で殆ど検出されないことから、この制御領域は、未分節中胚葉におけるtbx6遺伝子の発現制御において中心的な役割を担うことが明らかとなりました。これらの結果から、tbx6遺伝子の転写は、TR1を介してTbx6タンパク質自身によって維持されていることが分かりました。先述したように、体節が形成される際にRipplyによってTbx6タンパク質が分解されますが、今回の研究で見出されたtbx6遺伝子の転写が自身によって維持されるという仕組みは、RipplyによってTbx6タンパク質が分解されると同時に、tbx6遺伝子の転写もスイッチがONからOFFに切り替わると考えられます。その結果、迅速にTbx6タンパク質とその供給源となるtbx6遺伝子の発現の双方が抑制されることで、効率的に次の体節境界の位置へ移行させる仕組みが備わっていると考えられます(図4)。
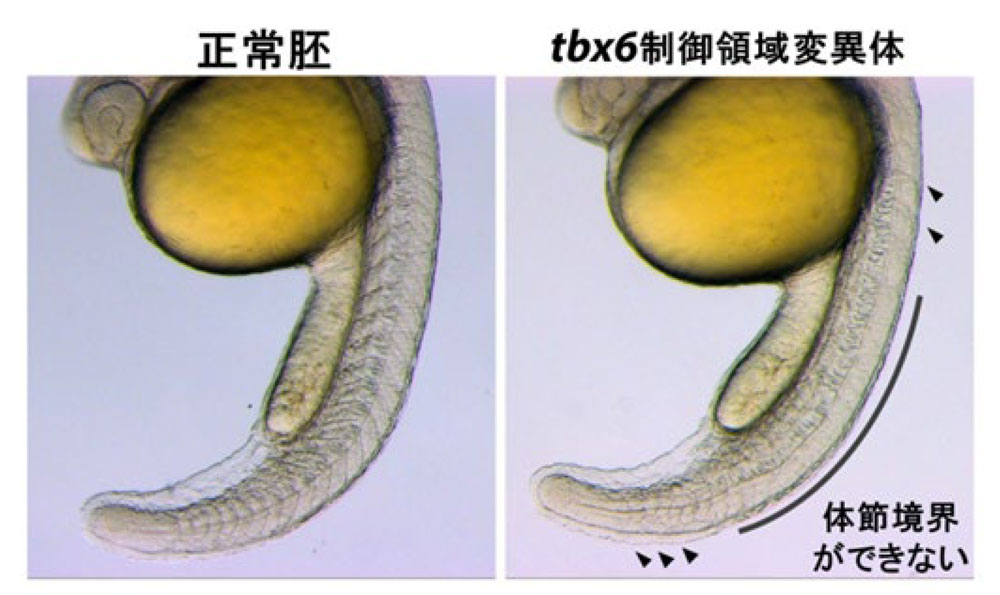 図3 tbx6制御領域の機能を欠失した変異体では、頭部側と尾部側の体節境界が一部形成されるが(矢尻)、7~25番目の体節境界が形成されない。
図3 tbx6制御領域の機能を欠失した変異体では、頭部側と尾部側の体節境界が一部形成されるが(矢尻)、7~25番目の体節境界が形成されない。
 図4 tbx6の転写制御は、Tbx6タンパク質がTR1領域を介して、自身の転写を活性化することにより維持される。RipplyによりTbx6タンパク質が除去されるが、それは同時にtbx6の転写が解除されることに繋がり、迅速にTbx6タンパク質の前方境界が後方へシフトすることを可能にしていると考えられる。
図4 tbx6の転写制御は、Tbx6タンパク質がTR1領域を介して、自身の転写を活性化することにより維持される。RipplyによりTbx6タンパク質が除去されるが、それは同時にtbx6の転写が解除されることに繋がり、迅速にTbx6タンパク質の前方境界が後方へシフトすることを可能にしていると考えられる。
【論文情報】
掲載誌 Development
論文名 Transcriptional autoregulation of zebrafish tbx6 is required for somite segmentation
著者名 伴 博之*, 横田大佑*, 乙坂栞里, 菊地守道, 木下宏史, 藤野友梨, 矢部泰二郎, 小原弘幹, 猪塚彩花, 赤間 燿, 弥益 恭, 高田慎治, 川村哲規
(* these authors contributed equally to this work)
【研究に関する問合せ先】
埼玉大学大学院理工学研究科 生命科学系専攻 生体制御学コース
川村哲規 准教授
")
