
小脳プルキンエ細胞の樹状突起では、樹状突起の興奮性が入力の伝搬を決める
2019-11-26 京都大学
大槻元 白眉センター特定准教授は、小脳の主要な出力細胞であるプルキンエ細胞の樹状突起では、樹状突起のそれぞれの興奮性が異なることによって、シナプス後電流が細胞体にまで伝導されるか否かが調節されることを見出しました。さらに、「可塑性」と呼ばれる神経細胞の興奮性が変化するメカニズムによって、シナプス後電流の伝わりやすさが変化することが分かりました。
近年、小脳の学習機能のメカニズムとして可塑性が注目され、これまでに「シナプス可塑性」(シナプス単位の電気活動が変化する現象)と「興奮性可塑性」(神経細胞レベルで活動電位発火が変化する現象)の2種類の可塑性の研究が進んできました。本研究では、プルキンエ細胞の細胞体と樹状突起から同時にシナプス後電流を記録することで樹状突起の興奮性可塑性の役割を調べました。その結果、興奮性可塑性が誘導されたり、カルシウム活性型カリウムチャネルの活性が抑えられた条件では、細胞体から遠い樹状突起(遠位樹状突起)へのシナプス入力が、効率よく細胞体まで伝導することが確認できました。さらに、同時記録したシナプス後電流の比を取る計算手法によって、プルキンエ細胞には平均4.5個のクラスター入力があり、その数は興奮性可塑性とカルシウム活性型カリウムチャネルの抑制によって減少することが分かりました。これは、樹状突起毎に伝わりやすさが異なっていたシナプス入力が、興奮性可塑性が誘導されることによって全体として細胞体まで通過しやすくなったことを意味します。
本研究成果は、小脳にはこれまで知られていなかった興奮性可塑性によるシナプス入力伝導の調節メカニズムが存在することを実証するもので、従来のシナプス可塑性に依存した小脳の学習理論との関わりをさらに検討することが今後の課題です。
本研究成果は、2019年11月22日に、国際学術誌「Journal of Neuroscience」のオンライン版に掲載されました。

図:本研究の概要図
詳しい研究内容について
小脳神経細胞の樹状突起で新規の学習原理を発見
―小脳プルキンエ細胞の樹状突起では、樹状突起の興奮性が入力の伝搬を決める!―
概要
京都大学白眉センター 大槻元 特定准教授は単独で、小脳の主要な出力細胞であるプルキンエ細胞(注1) の樹状突起 (注2)では、樹状突起のそれぞれの興奮性が異なることによって、シナプス後電流 (注3)が細胞体に まで伝導されるか否かが調節されることを見出しました。さらに、「可塑性」と呼ばれる神経細胞の興奮性が 変化するメカニズムによって、シナプス後電流の伝わりやすさが変化することが分かりました。
近年、小脳の学習機能のメカニズムとして可塑性が注目され、これまでに「シナプス可塑性」(シナプス単 位の電気活動が変化する現象)と 「興奮性可塑性」 (神経細胞レベルで活動電位発火が変化する現象) (注 4)の 2 種類の可塑性の研究が進んできました。本研究では、プルキンエ細胞の細胞体と樹状突起から同時にシナプス 後電流を記録することで樹状突起の興奮性可塑性の役割を調べました。その結果、興奮性可塑性が誘導された り、カルシウム活性型カリウムチャネル (注 5)の活性が抑えられた条件では、細胞体から遠い樹状突起 (遠位樹 状突起)へのシナプス入力が、効率よく細胞体まで伝導することが確認できました。さらに、同時記録したシ ナプス後電流の比を取る計算手法によって、プルキンエ細胞には平均 4.5 個のクラスター入力 (注 6)があり、そ の数は興奮性可塑性とカルシウム活性型カリウムチャネルの抑制によって減少することが分かりました。これ は、樹状突起毎に伝わりやすさが異なっていたシナプス入力が、興奮性可塑性が誘導されることによって全体 として細胞体まで通過しやすくなったことを意味します。以上の研究成果は、小脳にはこれまで知られていな かった興奮性可塑性によるシナプス入力伝導の調節メカニズムが存在することを実証するもので、従来のシナ プス可塑性に依存した小脳の学習理論との関わりをさらに検討しなくてはなりません。
本研究成果は、2019 年 11 月 22 日に、国際学術誌「Journal of Neuroscience」のオンライン版に掲載され ました(単独 筆頭・責任著者としての論文)。
1.背景
小脳は哺乳類の脳後部に位置し、動物の運動やバランスに重要な働きを持つ脳部位です。また、運動学習と 呼ばれる無意識の反射運動の学習過程にもかかわります。近年では、小脳は単に運動機能だけにかかわるので はなく、様々な脳部位と協調して活動することで、発音 ・発話、体性局在、課題遂行、注意など、様々な機能 を持つ高次な脳機能を担う部分であることがわかり始めています。このような小脳の学習機能のメカニズムと して、最も注目を浴びている現象の一つは「可塑性」と呼ばれるものです。「可塑性」は脳の神経細胞が外部 からの刺激に適応して変化する性質であり、動物が特殊な刺激を受けたり、繰り返しの動作があったりしたと きに起きる神経活動によって誘導されます。「可塑性」が神経細胞に誘導されると、神経細胞の興奮を伝える シナプス伝達が大きくなったり、神経細胞の発火頻度 (注 7)が増加したりする状態が続きます。これまでに研究 が進んでいるタイプの可塑性は、シナプス可塑性 (シナプス単位の電気活動が変化する現象)と興奮性可塑性 (神経細胞レベルで活動電位発火が変化する現象)です。
従来、小脳学習における基盤原理はシナプス可塑性であると多くの研究者が支持してきました。特に、プル キンエ細胞に形成される平行線維投射 (平行線維は小脳顆粒細胞から伸びて興奮を伝達する神経線維)におけ るシナプス可塑性異常と、動物の運動学習異常との間に強い相関がみられることから、20 年以上に渡ってこ の仮説は支持されてきました。しかしながら、小脳神経回路では、平行線維のシナプスだけではなく、多くの 種類の神経細胞間でもシナプス可塑性が誘導され、小脳依存的学習に関与することも分かり始めています。
一方、小脳プルキンエ細胞の興奮性可塑性は、2010 年からその発現や誘導メカニズム、運動学習への寄与 が次第に明らかにされてきました (Schonewill et al., 2010, Neuron; Belmeguenai et al., 2010, J Neurosci.)。 大槻特定准教授らは本研究に先立って、プルキンエ細胞の樹状突起と細胞体からの電流を同時に長時間記録す ることに成功し、プルキンエ細胞に興奮性可塑性が誘導されたときに、その樹状突起ごとの興奮性も上がるこ とを示しました。また、このような樹状突起での興奮性増大には、SK チャネル(カルシウムイオン(Ca2+) 活性化型カリウムイオン(K +)チャネル)の機能低下が関わっていることも明らかにしていました(Ohtsuki et al., 2012, Neuron, Ohtsuki & Hansel, 2018, iScience)。しかし、樹状突起からの電気記録自体が非常に難 しく、挑戦する研究室の数も非常に少ない状況であったため、樹状突起のシナプス伝達がどのように伝導され るのかについては、モデルによるシミュレーションによって予想されるのみで、実際のメカニズムは全く分か っていませんでした。そこで本研究は、上記の先行研究を踏まえて、樹状突起のシナプス伝達が細胞体まで到 達する過程に変化が見られると予想し、そのメカニズムを解明するとともに、小脳の学習原理を再考すること を目指して遂行されました。
2.研究手法・成果
本研究では、小脳プルキンエ細胞における細胞レベルでの学習メカニズムを再考するために、プルキンエ細 胞の樹状突起と細胞体からシナプス後電流を同時に記録して、興奮性可塑性が誘導されるとどのようにシナプ ス後電流の伝導過程に変化が見られるのかを調べました。本研究ではおもに、「パッチクランプ法」と呼ばれ る、微細なガラス管を使って一つ一つの神経細胞の電気活動を直接計測する電気生理学的な手法を用いました。 プルキンエ細胞の樹状突起と細胞体から同時に電気記録する実験には、高度な実験技術を要します。この手法 に加えて、シミュレーションモデルを使って、実験によって得られたデータを統計学的に検証しました。その 結果解明された主なポイントは以下の 2 つです。(次ページ図参照)
1. プルキンエ細胞の遠位樹状突起 (細胞体から遠くに位置する樹状突起)に入力されるシナプス後電流 は、興奮性可塑性の誘導、SK チャネルの阻害、K +チャネルの広範囲な阻害によって、細胞体にまで 通過しやすくなることが判明した。
2. 細胞体と樹状突起から記録されるシナプス後電流の比を調べることで、平均 4.5 個のクラスター入力 が推定された。また、このクラスター入力の数は、興奮性可塑性の誘導と SK チャネルの阻害によっ て減少することが分かった。
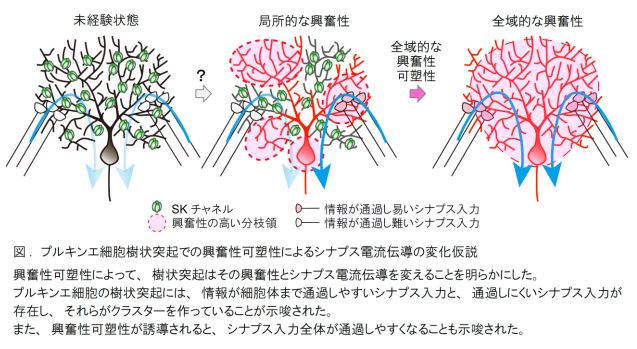
これらの結果は、小脳プルキンエ細胞の樹状突起に入力されるシナプス伝達は、シナプス可塑性とは異なる、 興奮性可塑性を介するメカニズムによって、細胞体にまで到達するか否かが決定されていることを意味します。 すなわち、プルキンエ細胞の樹状突起には、興奮性の増大と SK チャネルの機能低下によって情報が細胞体ま で通過しやすいシナプス入力と、通過しにくいシナプス入力が存在し、それぞれが集中するクラスターを作っ ていることが示唆されました。また、プルキンエ細胞全域に興奮性可塑性が誘導されると、シナプス入力全体 が細胞体まで通過しやすくなることも示唆されました。
従来は、細胞内液にセシウムイオン(Cs+)を含んだ溶液を使用した実験が多かったために、これらの現象 は全く見出すことは出来ませんでした。本研究成果は、これまでに考えられていたシナプス可塑性に基づく小 脳の学習理論に、興奮性可塑性という異なった側面から樹状突起レベルでの学習メカニズムを提案するもので、 これまでの細胞レベルでの学習メカニズムに対する考え方の再考と再検証を促す重要な結果です。
3.波及効果、今後の予定
上述のように、本研究では、小脳プルキンエ細胞における樹状突起レベルでの学習メカニズムを解明しまし た。小脳だけではなく、脳全体の神経細胞レベルでの学習メカニズムに対する従来説に再考を促す結果です。
しかしながら、本研究ではラットの小脳切片だけを使用しており、実際に生きた脳から学習中に電気記録す ることは出来ていません。電気活動イメージングなどの手法によって、これらの結果は検証することが可能で す。また、げっ歯類以外の動物 (霊長類などの大型哺乳類や、爬虫類 ・魚類などのより原始的な動物)でも同 様の結果が得られるのか否かについても、さらなる検証が必要です。
最近の大槻特定准教授らの研究結果によって、小脳プルキンエ細胞における興奮性可塑性は、鬱様症状など の動物の精神疾患様行動異常と関連があることが分かりました(Yamamoto et al., 2019, Cell Reports)。プル キンエ細胞の興奮性可塑性は、小脳の急性炎症時に、ミクログリアと呼ばれる脳内の免疫細胞の異常な活動に よって誘導されます。したがって、微生物感染後の急性炎症時に、樹状突起レベルでの学習メカニズムに異常 が起こることも予想されます。たとえば、樹状突起毎に重要な情報と、そうでない情報をより分けることがで きないことで、精神状態に影響を与えるのかもしれません。この点についても、今後の詳細な研究が必要です。
4.研究プロジェクトについて
本研究プロジェクトは、京都大学白眉センター所属の研究者 1 名 (大槻特定准教授)によって遂行され、京 都大学白眉運営費、興和生命科学振興財団、日本学術振興会科学研究費助成事業(科研費)、内藤記念科学振 興財団、ブレインサイエンス振興財団の助成を受けました。
<用語解説>
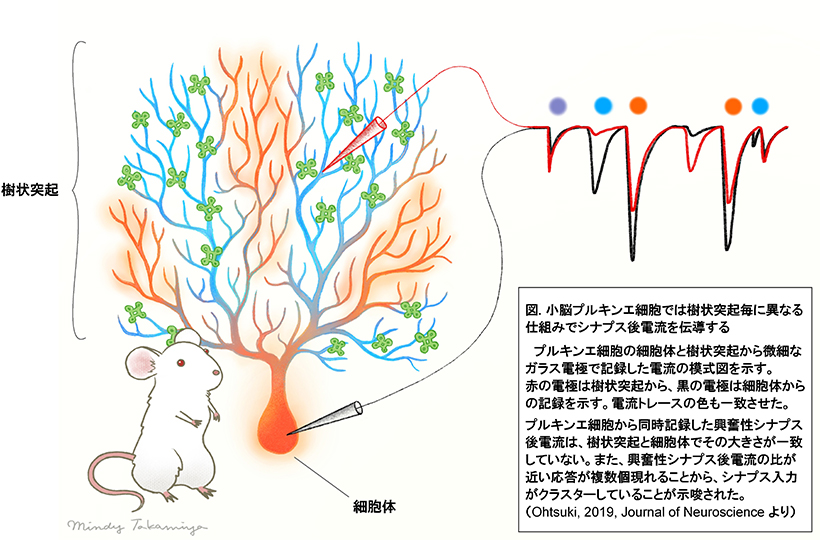
(注1)プルキンエ細胞: 小脳の主要な神経細胞 (1 ページの図参照)。プルキンエ細胞は、顆粒細胞など他 の神経細胞からの入力を受けて、小脳核へと抑制性の出力を行う。小脳皮質の主要な情報処理を行うと考 えられている。
(注2)樹状突起: 神経細胞は基本形態として、樹状突起、細胞体、軸索から成り立つ。樹状突起にはスパイ ン (突起棘)と呼ばれる極小の棘 (1マイクロメートル程度の大きさ)が数千〜数十万個あり、そこに他 の神経細胞との接続部位が形成される。これをシナプスと呼ぶ。神経細胞のシナプス入力構造である樹状 突起は、まさに「樹の枝」のようにその枝幹を広げて、多数のシナプスを形成する。遠位樹状突起とは、 枝の末端の部位のこと。樹状突起に入力された電機情報が細胞体で集約されて、活動電位が発生する (注 5 を参照して下さい)。
(注3)シナプス後電流: 伝達物質を受けた後にシナプス後部に流れる電流のこと (1 ページの図中のトレー スを参照)。特に電位固定下で観察できる。一般に、活動電位が軸索の末端まで伝搬されると、そこに形 成されるシナプス前終末を活性化させる。すると、シナプス小胞が開いて、蓄えられていた伝達物質が放 出される。このシナプス後電流が樹状突起で集約されることで、活動電位発生が決まる。
(注 4)興奮性可塑性: 神経細胞の発火頻度が長時間増大する現象である。英語では、Intrinsic Plasticity と呼 ばれる。この興奮性可塑性は、シナプス伝達効率が長期間増大するシナプス可塑性 (長期増強)とは異な り、カルシウム活性型カリウムチャネルの下方調節が関わる。カルシウム活性型カリウムチャネルの活性 が下がると、神経細胞の膜特性は分極(負の電荷を発生させること)しにくいので、興奮しやすくなる。
(注 5)カルシウム活性型カリウムチャネル: 生体組織内の Ca2+イオンに反応して開く K +イオンチャンネル。 K +イオンチャンネルには様々なタイプが存在して、神経細胞では細胞膜を分極させる役割を持つ。カルシ ウム活性型カリウムチャネルの場合は、たとえば、Ca2+イオン濃度上昇をともなう神経活動があった場合 に活性化されて、神経細胞を局所的に分極させる。
(注 6)クラスター入力: シナプス入力が集中して一部の樹状突起などに入力すること。生体内ではシナプス 後電位のホットスポットとして観察されると考えられているが、その詳細は明らかではない。
(注 7)神経細胞の発火: 神経細胞はその活動として、活動電位と呼ばれる 100mV、1ミリ秒程度の微弱な電 気パルスを、おもに細胞体の付け根である軸索起始部付近で発生させる。この電気パルスを活動電位と呼 び、その発火頻度によって次の細胞に伝える情報が変化する。
<研究者のコメント>
本プロジェクトは小脳の学習メカニズムについて、樹状突起の興奮 性可塑性がどのように シナプス入力を制御するのか調べることを目 的としたものです。樹状突起からの電気生理学的手法は難易度の高い 実験手法ですが、効率的にデータを積み重ねることが出来ました。か つての留学先であるシカゴ大学医学部での研究結果を再検討するこ とが、このメカニズムに気付くために、非常に役に立ちました。日本 に居ただけでは、全く得られなかった成果です。他の研究機関の生理 学分野の研究者の方々にも、多くの興味を持って受け入れていただい ております。
また、独立研究者として白眉センターに研究期間を頂けたおかげで、従来の定説にとらわれることなく、自 由な発想で新たな研究領域を開くことができたと思います。今回は単独研究を行いましたが、多くの研究者 (学 部生・大学院生)に開かれるべきチャンスだったと思います。京都大学として、共に頑張りましょう。
<論文タイトルと著者>
タイトル:Modification of synaptic-input clustering by intrinsic excitability plasticity on cerebellar Purkinje cell dendrites (小脳プルキンエ細胞の樹状突起における興奮性可塑性がシナプス入力のクラスタリ ングを変化させる)
著 者:大槻 元
掲 載 誌:Journal of Neuroscience DOI:10.1523/JNEUROSCI.3211-18.2019

