2020-02-04 甲南大学,日本医療研究開発機構
甲南大学大学院自然科学研究科の久原篤 教授、高垣菜式 博士後期課程院生、太田茜 研究員らの研究チームは、触った時の感触といった機械刺激注1)を受容する「メカノ受容体注2)」が「体の低温耐性」を調節していることを線虫の解析から明らかにしました。
温度は最も重要な環境情報の一つであり、生命にとって温度変化への適応は必須です。研究チームはシンプルなモデル実験動物である線虫C.エレガンス注3)の低温耐性注4)を解析してきました。今回、ヒトではプリン体の代謝に関わることが知られているキサンチン脱水素酵素(XDH)注5)を欠損した線虫において低温耐性の喪失が見られました。XDHは線虫頭部のAINとAVJ注6)と呼ばれる各一対の介在ニューロン注7)で低温耐性を調節していました。AINとAVJは介在ニューロンであるため、これらの2つの介在ニューロンの上流には未知の温度受容ニューロンがあると仮説を立てました。
この仮説に基づき解析を進めたところ、新たな温度受容ニューロンASG注8)が見つかりました。ASGの温度受容には、機械刺激を受容するDEG/ENaC型注9)のメカノ受容体であるDEG-1が関わっていました。さらに、DEG-1を味覚ニューロンASE注10)に導入したところ、味覚ニューロンASEが温度に反応するようになりました。同様に、DEG-1をアフリカツメガエルの卵母細胞注11)に導入すると、温度に反応するようになりました。さらに興味深いことに、線虫のDEG-1に相当する「ヒトのMDEG注12)」をアフリカツメガエルの卵母細胞に導入したところ、温度に反応するようになりました。
以上の研究から、DEG/ENaC型のメカノ受容体が線虫の低温耐性に関わることが見つかりました。これは、ヒトを含む動物においてメカノ受容体DEG/ENaCが体の温度適応に関わることを示す初めてのケースです。
さらに、ヒトのMDEGも温度に反応したことから、ヒトの温度受容の仕組みや、温度による健康障害の原因解明などに役立つと期待されます。
本研究成果は、ドイツ標準時2020年2月3日(日本時間:2月3日)に、欧州分子生物学機構誌「EMBO reports(エンボ リポーツ)」のオンライン速報版で公開されました。
研究の背景と経緯
温度は、生体反応に直接影響を及ぼす重要な環境因子の一つです。そのため、生物が生存していくためには、温度変化に対して適切に応答することが重要です。例えば、ヒトでは体温がわずか3℃低下し33℃になると仮死状態になることや、病気の発生率や死亡率は気温変化が大きい時期に増えます。温度が関わる病気は多数あり、急激な温度変化によるヒートショック死や低体温症が社会問題となっており、これらにより亡くなられる方は交通事故による死亡数よりも多いという報告があります。地球規模でも、温暖化や局所寒冷化により生物種の3割が絶滅の危機になると報告されており、生物の温度適応は世界レベルの課題となっています。
このように、生物の温度応答や温度適応の解明は多方面から望まれていますが、一方で、ヒトの温度応答は体内の多数の細胞からなるネットワークで制御されています。例えば、ヒトの体は約37兆個の細胞で構成されており、生体調節の中枢である脳は、約1000億個の神経細胞で構成されています。そのため、その細胞間のネットワークは非常に複雑であり、ヒトの生体内の情報処理を解析する大きなハードルになっています。
本研究チームは、959個の細胞からなるシンプルなモデル実験動物である線虫「C.エレガンス」を使い、温度応答のメカニズムの解明を目指してきました。これまでに、低温耐性現象(図1)の解析から、低温耐性に関わる分子と組織のネットワークが少しずつ見つかってきました。具体的には、頭部に一対ずつ存在するASJとADLと呼ばれる感覚ニューロンがそれぞれ温度を受容する温度受容ニューロンであること、ASJからインスリンが分泌され、腸がインスリンを受容することによって体内の脂肪酸注13)の構成比を変化させ、低温耐性を低下させることを見つけました(Ohta et al., Nature commun., 2014; Ujisawa., PNAS, 2018; Okahata., Science Advances, 2019)。更に、腸から精子に情報が伝達され、精子が頭部の温度受容ニューロンをフィードバック制御注14)することを見つけました(Sonoda et al., Cell reports, 2016)。
このように低温耐性を低下させる仕組みは明らかになってきていましたが、低温耐性を増強させる仕組みはほとんど解析されていませんでした。

図1:線虫の低温耐性線虫は、25℃で飼育された後に2℃に置かれると死滅するが、15℃で飼育された後に2℃に置かれると生存できる(低温耐性現象)。
研究の内容
キサンチン脱水素酵素XDH-1は2つの介在ニューロンを介して低温耐性を制御
低温耐性に関わる新しい遺伝子を見つけるために、飼育温度に依存して発現量を変動させる遺伝子を多数解析し、ヒトにおいてプリン体の代謝に関わる遺伝子として知られているキサンチン脱水素酵素に相当する、線虫のキサンチン脱水素酵素であるXDH-1を発見しました。
XDH-1が欠損した変異体線虫(xdh-1変異体)は、15℃で飼育後に2℃の低温刺激を与えると野生株に比べて低い生存率を示す異常が見られました(図2)。さらに興味深いことに、このxdh-1変異体の低温耐性異常は、AINやAVJと呼ばれるわずか2つの介在ニューロンに正常なxdh-1遺伝子を導入すると回復することが分かりました。つまり、XDH-1はAINとAVJの2つの介在ニューロンで働くことで、低温耐性を制御することが示唆されました。
XDH-1のAINとAVJでの働きが低温耐性に必要であると判明したため、カルシウムイメージング法注15)という光技術で、xdh-1変異体のAINとAVJそれぞれの温度変化に対する神経活動の変化を調べました。解析の結果、XDH-1を欠損させた変異体ではAINの神経活動が低下する異常が見られ、AVJでは反対に神経活動が上昇する異常が見られました。これらの異常な神経活性はxdh-1変異体のAINとAVJそれぞれに正常なxdh-1遺伝子を導入すると回復しました。つまり、XDH-1が2つの介在ニューロン(AINとAVJ)の神経活動を制御していることが示唆されました(図3)。
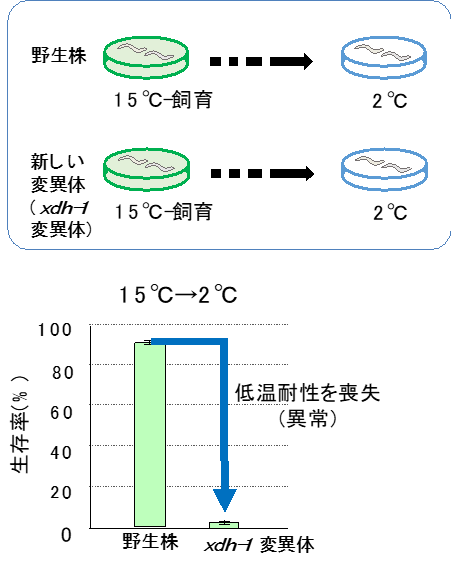
図2:xdh-1変異体は低温耐性を喪失している野生株は通常、15℃飼育後に2℃に置かれても生存できるが、キサンチンデヒドロゲナーゼが欠損しているxdh-1変異体は15℃飼育後に2℃の低温刺激にさらされると死滅する。
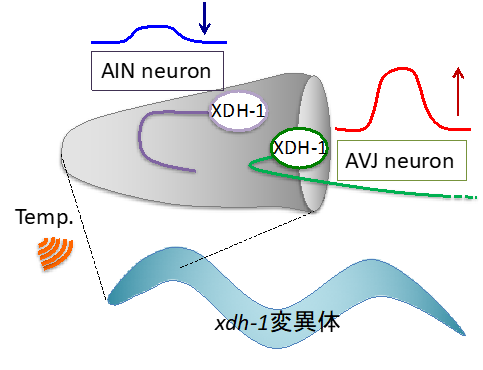
図3:XDH-1欠損が神経の活動を左右するxdh-1変異体のAINニューロンは反応が低下する異常を示し、AVJニューロンは過活性化する異常を示す。これらの異常がxdh-1変異体の低温耐性異常を引き起こしていると考えられる。
DEG/ENaC型メカノ受容体DEG-1を含む低温耐性の新たな神経回路
AINとAVJ は神経情報を橋渡しする介在ニューロンであるため、これらの介在ニューロンの上流には温度を受容する感覚ニューロンが存在する可能性が考えられました。AINとAVJの上流には9つの感覚ニューロンが存在し、そのうちの5つの感覚ニューロンにおいて、機械刺激を受容するDEG/ENaC型のメカノ受容体が発現していました。そこで、これらのメカノ受容体を発現する感覚ニューロンが低温耐性に関与するのではないかと考え、メカノ受容体の解析を行いました。すると、DEG/ENaC型のメカノ受容体であるDEG-1の変異体において、低温耐性の異常が見つかりました。さらにDEG-1を発現するASG感覚ニューロンが温度に応答し、その温度応答はdeg-1変異体において低下していました。この異常な反応の低下はdeg-1変異体のASG感覚ニューロンに正常なdeg-1遺伝子を導入することで回復しました。つまり、メカノ受容体であるDEG-1がASGの温度応答に関与していました。
ASG感覚ニューロンはAIN介在ニューロンの上流に位置していたことから、DEG-1のASG感覚ニューロンにおける働きの結果として、AINとAVJ介在ニューロンの活性が変化している可能性を考えました。そこで、メカノ受容体DEG-1の変異体におけるAINとAVJ介在ニューロンの温度への応答を調べました。
カルシウムイメージングによる解析から、メカノ受容体DEG-1が異常になるとAINとAVJ介在ニューロンの温度に対しする反応が異常になることが分かりました。そこで、deg-1変異体のASG感覚ニューロンにのみ正常なdeg-1遺伝子を導入すると、AINとAVJ介在ニューロンの異常な神経活動が回復しました。つまり、メカノ受容体DEG-1のASG感覚ニューロンにおける働きの結果として、AINとAVJの2つの介在ニューロンの活性が変化していると考えられました。
メカノ受容体DEG-1を介した温度応答
DEG-1がASG感覚ニューロンの温度応答に関わっていたことから、メカノ受容体であるDEG-1が温度を受容しているという仮説を立てました。この仮説を検証するために、温度上昇には反応しない味覚ニューロン(ASE)にDEG-1を強制発現注16)させ、温度に応答するようになるかを解析しました。カルシウムイメージングによる解析の結果、DEG-1を強制発現させたASEは温度上昇に反応を示すようになりました。
DEG-1が温度を受容するかを直接的に示すために、アフリカツメガエルの卵母細胞を用いた電気生理学的解析注17)を行いました。通常、卵母細胞は温度変化に反応を示しませんが、DEG-1を強制発現させた卵母細胞は温度に対して電流値の変化を示すようになりました(図4)。さらに、線虫のDEG-1に相当する「ヒトのMDEG」を強制発現させた卵母細胞も温度に反応するようになりました。
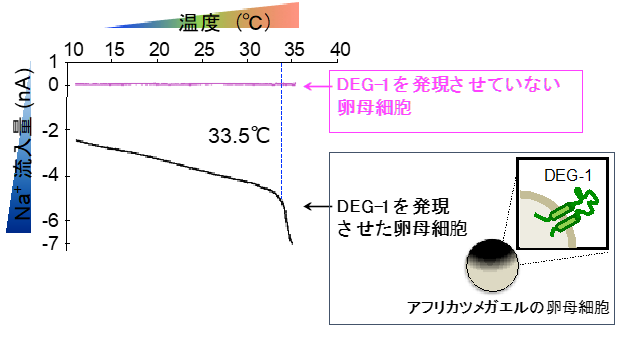
図4:アフリカツメガエル卵母細胞へのDEG-1強制発現の解析DEG-1をアフリカツメガエルの卵母細胞に強制発現させ、温度に対する反応を測定した。DEG-1を強制発現させた卵母細胞は温度に対してNa+の流入を示した。
今後の展開
本研究から、線虫からヒトまで保存されている「DEG/ENaC型のメカノ受容体」が細胞の温度応答に関わり、線虫では体の低温耐性に関わることが見つかりました。この結果は、動物においてメカノ受容体DEG/ENaCが個体の温度適応に関わることを示す初めてのケースです。
線虫では、メカノ受容体DEG-1を介してASG感覚ニューロンが受け取った温度情報が、キサンチン脱水素酵素XDH-1が機能するAINとAVJの2つの介在ニューロンを介し、「体の低温耐性を変化」させていると考えられます(図5)。一方で、低温耐性におけるキサンチン脱水素酵素(XDH)およびキサンチンの役割については、未知の点が残されています。
DEG-1やヒトのMDEGは機械刺激を受容するメカノ受容体であることから、温度そのものを受容している可能性に加え、温度変化による細胞膜の流動性の変化などを受容している可能性も考えられます。今後のDEG/ENaCを介した温度応答の仕組みの解析から、温度受容体が温度をどのように感じているのかを解き明かす鍵が見つかってくることを期待しています。
ヒトを含む高等動物においても、DEG/ENaC型のメカノ受容体が機械刺激の受容や、酸味や塩味の受容に機能していることが報告されています。更に、本研究から、ヒトのMDEGも温度受容に関わることが示唆されました。そのため、ヒトにおける温度応答の分子メカニズムの解明と、温度が関わる未解明の疾患の原因解明に繋がると期待されます。
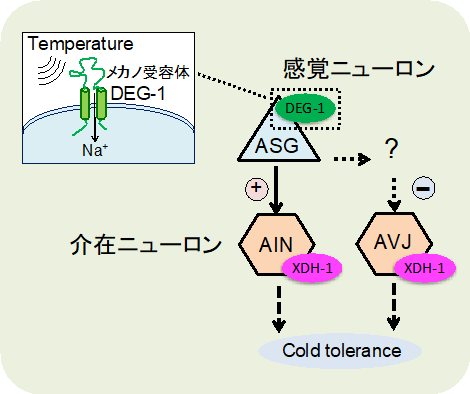
図5:低温耐性を正に制御する新たな神経回路モデルDEG-1が温度を受容するとASG感覚ニューロンが反応し、AIN介在ニューロンを活性化させ(矢印)、AVJ介在ニューロンを抑制する(点線矢印)。その結果として、体の低温への耐性が獲得される。個体の温度応答に関わる新たな神経回路が見つかった。
研究支援
本研究は、日本医療研究開発機構(AMED)革新的先端研究開発支援事業(PRIME)「メカノバイオロジー機構の解明による革新的医療機器及び医療技術の創出」研究開発領域における研究開発課題「新規温度センサー分子の単離とそれを用いた応用技術の創出」(研究開発代表者:久原篤)、文部科学省科学研究補助金 新学術領域(研究領域提案型)「温度を基軸とした生命現象の統合的理解」研究領域における研究課題「TRPチャネルおよび膜脂質による温度センシング機構の解明」(研究代表者:富永真琴、研究分担者:久原篤)の研究費によって行われました。
用語解説
- 注1)機械刺激
- 機械刺激とは物理的な力による刺激などを指す。皮膚への接触刺激やヒトの内耳の鼓膜への振動刺激などが含まれる。
- 注2)メカノ受容体
- 動物において、皮膚や神経系などに存在する機械刺激を受け取るタンパク質などを指す。皮膚などへの機械刺激によって生じる細胞膜の張力を感じて開き、細胞内にイオンを透過させるイオンチャネル型のメカノ受容体などが知られている。
- 注3)C.エレガンス
- 土壌に生息する非寄生性の線虫であり、正式名称はカエノラブディティス・エレガンス。古くから分子遺伝学的な解析に使用されている。1998年には多細胞生物で初めて全ゲノムDNAの解読が終了した。ヒトの遺伝子数と同等に約2万個の遺伝子を持ち、類似の遺伝子も多い。
- 注4)低温耐性
- 動植物が寒期を乗り越える際に、秋の低くなってきた温度を感じ取り、到来する冬に向けて生体内で低温への耐性を構築する現象。
- 注5)キサンチン脱水素酵素(XDH)
- ヒトでは、プリン体の代謝経路でヒポキサンチンからキサンチン、キサンチンから尿酸への代謝を触媒している。
- 注6)AINとAVJ
- C.エレガンスの頭部にそれぞれ左右一対存在し、感覚ニューロンが受け取った情報を橋渡しする介在ニューロンである。本研究から、これらのニューロンが低温耐性に関わることが分かってきた。
- 注7)介在ニューロン
- ニューロン間の情報伝達を橋渡しするニューロン。
- 注8)ASG(温度受容ニューロン)
- C.エレガンスの頭部に左右一対だけ存在し、環境の感覚情報を受け取る感覚ニューロンであり、線虫が好む水溶性誘引物質を感じる。本研究から、このニューロンが温度を感じる温度受容ニューロンであることが分かってきた。
- 注9)DEG/ENaC型のメカノ受容体
- Degenerin/Epithelial Sodium Channelの略称。機械刺激などを受容する3量体のメカノレセプター(機械受容体)であり、Na+を流入する。本研究から、線虫のDEG/ENaC型メカノ受容体が低温耐性において温度を受容する可能性が得られた。
- 注10)ASE味覚ニューロン
- C.エレガンスの頭部に左右一対だけ存在し、環境の感覚情報を受け取る感覚ニューロンである。主に塩味(NaCl)を受容し、C.エレガンスが適度な濃度のNaClに集まる行動に関わっている。
- 注11)アフリカツメガエルの卵母細胞
- 電気生理学的な解析に使用される未受精卵。さまざまな種類の生物のチャネルや受容体を発現させ、さまざまな刺激への反応を測定できる。
- 注12)ヒトのMDEG
- 線虫DEG-1のヒトでのホモログ(相同体:遺伝子や形態が共通の祖先に由来)。低温下で反応が増強されることが報告されているが、温度を受容するかは分かっていなかった。最近の研究から、温度を受容する可能性が得られていた。
- 注13)脂肪酸
- 広義には油脂や脂質などの構成成分である有機酸を指す。生物の低温下での生存の観点においては、脂肪酸の固まりやすさである飽和度が重要と言われている。不飽和脂肪酸は、低温度でも固まりにくい「油」であるため、低温下において変温動物は、生体膜中の不飽和脂肪酸比率を増やし、低温下でも生存できるようになっていると考えられている。
- 注14)フィードバック制御
- 生体が環境に適応するために、体内の環境を調節する仕組みのひとつである。生体に環境情報の入力が入った場合に、その入力刺激が大きくなり過ぎたり小さくなり過ぎたりしないように、出力に近い末梢の細胞から入力に近い上位の細胞を調節する仕組みのひとつ。フィードバック制御には、上位の細胞をさらに活発に働かせるための「正のフィードバック制御」と上位の細胞の活動を抑えるための「負のフィードバック制御」が存在する。
- 注15)カルシウムイメージング法
- ニューロンの働きに重要である細胞内のカルシウム濃度の変動を色の変化として見る方法のことを指す。本研究では、カルシウムイオンと結合し、カルシウムの濃度により色が変わる蛍光タンパク質(カメレオンやGCaMP)を細胞に導入した。
- 注16)強制発現
- 普段は発現していない遺伝子を人工的に細胞内で発現させること。今回、特定の細胞内において、本来はほとんど存在しない種類のタンパク質を、遺伝子導入により強制的に作らせた。
- 注17)電気生理学的解析
- 神経細胞や筋肉細胞などの電気的性質と生理的な働きとの関係を調べるための解析手法を指す。本研究においては、細胞内のイオンチャネルや受容体の特性を、環境刺激を与えた際の細胞内に生じる電流や電圧の変化として直接的に測定した。
掲載論文名
“The mechanoreceptor DEG-1 regulates cold tolerance in Caenorhabditis elegans”
(メカノ受容体を介した温度受容がC.エレガンスの低温耐性を制御する)
EMBO reports, 2020, e48671(DOI: 10.15252/embr.20194867)
発表者
*責任著者
高垣 菜式(タカガキ ナツネ)(甲南大学 博士後期課程生、日本学術振興会特別研究員DC2)
*太田 茜(オオタ アカネ)(日本学術振興会特別研究員RPD、甲南大学特別研究員)
大西 康平(オオニシ コウヘイ)(甲南大学 博士後期課程生、日本学術振興会特別研究員DC2)
川鍋 陽(カワナベ アキラ)(香川大学医学部医学系研究科 研究員)
水口 洋平(ミナクチ ヨウヘイ)(国立遺伝学研究所、研究員)
豊田 敦(トヨダ アツシ)(国立遺伝学研究所、特任教授)
藤原 祐一郎(フジワラ ユウイチロウ)(香川大学医学部医学系研究科 教授)
*久原 篤(クハラ アツシ)(甲南大学理工学部/大学院自然科学研究科/統合ニューロバイオロジー研究所、教授)
お問い合わせ先
本研究内容に関すること
高垣 菜式(タカガキ ナツネ)(本研究論文の筆頭著者)
日本学術振興会 特別研究員DC2
甲南大学大学院自然科学研究科博士後期課程生
久原 篤(クハラ アツシ)(本研究論文の責任著者)
甲南大学 理工学部 教授
太田 茜(オオタ アカネ)(本研究論文の責任著者)
日本学術振興会 特別研究員RPD
甲南大学特別研究員
広報に関すること
甲南学園 広報部
AMED事業に関すること
国立研究開発法人日本医療研究開発機構(AMED)
基盤研究事業部 研究企画課

