
ヒト大脳新皮質の進化過程を解き明かす
2020-07-10 慶應義塾大学医学部,実験動物中央研究所,理化学研究所
概要
マックスプランク分子細胞生物学遺伝学研究所のウィーランド・フットナー教授、ミヒャエル・ハイデ研究員、慶應義塾大学医学部生理学教室の岡野栄之教授(兼 理化学研究所脳神経科学研究センター〈CBS〉マーモセット神経構造研究チーム チームリーダー)、村山綾子特任助教(兼CBS客員研究員および実験動物中央研究所兼任研究員)、実験動物中央研究所の佐々木えりか部長、黒滝陽子室長、篠原晴香研究員のグループは以下の研究を行い、ヒトに特異的な脳の拡大の機序を解明しました。
まず、ヒトにしか存在しない遺伝子であるARHGAP11Bを、本来健常人で発現している生理的に近い量で発現する非ヒト霊長類であるコモンマーモセット(注1)(以下、ARHGAP11B遺伝子導入マーモセット)を作製しました。胎生後期の脳の解剖学的解析と神経系細胞の定量的解析を行ったところ、ARHGAP11B遺伝子導入マーモセットでは脳のシワ(脳回脳溝構造:注2)が本来ない場所に形成され、GI指数(脳の凹凸を示す指数)が野生型の約1.1倍に増高していました。これは、神経前駆細胞の一種であるbRG細胞(注3、図1-2)の増加に伴って、脳表面積の拡大に寄与している脳表近く(浅層)の神経細胞が約20%増加していることに起因することが分かりました。以上の結果から、今回、研究グループは、ARHGAP11Bはチンパンジーに至る進化系統から分岐したあとの、ヒトの進化過程における大脳新皮質(図1-1)の拡大と脳回脳溝構造の増加をもたらした原因遺伝子であることを明らかにしました。すなわち、ARHGAP11Bによってもたらされた大脳新皮質の拡大は、ヒトに特徴的な脳機能の獲得に関連していると考えられます。
本研究成果は、2020年6月18 日(中央ヨーロッパ時間CEST)、学術科学雑誌『Science』に掲載されました。
研究の背景
大脳新皮質の進化
知覚、記憶、言語、思考などといった高次の脳機能をつかさどるヒトの大脳新皮質(図1-1)は、最も近い近縁種であるチンパンジーの約3倍もの大きさがあり、その機能も特に発達しています。このように進化の過程で大きくなった脳が頭蓋骨の限られた空間に収まるために、脳は折りたたまれ、シワができたと考えられています。それでは、ヒトはどのようにして大きな新皮質と多くのシワを獲得してきたのでしょうか。

図1 大脳皮質の構造模式図(左)図1-1:大脳新皮質の位置(濃ピンク)
(右)図1-2:大脳皮質断面図(左の図1-1内の点線(黒)での断面図)
脳表側と脳室の両方向に長い突起を伸ばしている神経幹細胞(赤)、脳表側にだけ長い突起を伸ばしているbRG細胞(濃ピンク)、多極性の短い突起を持つ中間型神経前駆細胞(青)、神経細胞(灰)の分裂様式をそれぞれ同色の矢印で示した。
ヒト特異的遺伝子とマウスやフェレットでの解析
2010年、ウィーランド・フットナー教授の研究グループは、大脳新皮質の拡大には、脳を構成している神経細胞を生み出す神経幹細胞(注4、図1-2)と並び、bRG細胞(図1-2)が大きく寄与していることを見出しました。そこで、ヒトbRG細胞に特異的に発現している遺伝子を探索し、2015年にARHGAP11B遺伝子を同定しました。このヒト特異的遺伝子ARHGAP11Bをマウス胎仔脳に過剰に発現させると、bRG細胞の数が増えて脳が拡大し、通常は滑脳(注5)であるマウスの脳にもシワのような凹凸ができることを発見しました(Florio Met al., Science, 2015)。さらに、もともと脳にシワがあるフェレット(注6)でも大脳新皮質が拡大してシワが増えることがわかりました(Kalebic N et al., eLIFE, 2018)。
ヒトの進化過程におけるARHGAP11Bの出現
ヒトにしかない遺伝子であるARHGAP11 Bは、多くの動物種がもっているRHGAP11A遺伝子の部分的重複(注7)によって生じたとされています(図2)。この遺伝子重複は約500万年前、チンパンジーに至る進化系統と、ネアンデルタール人からヒトに至る進化系統が分岐した後に起こりました。そこからさらに長い年月を経た後、今から150万年~50万年前までの間に、ARHGAP11B遺伝子はたった1ヶ所の変異を生じました。その結果、ARHGAP11B はbRG細胞を増加させる能力、つまり大脳新皮質を拡大する機能を獲得したとフットナー教授らのグループは推定しました(Florio M et al., Science Advances, 2016)。しかし、ARHGAP11Bがヒトの大脳新皮質の進化過程に関与することを証明するためには、霊長類でのARHGAP11Bの機能解析が求められていました。
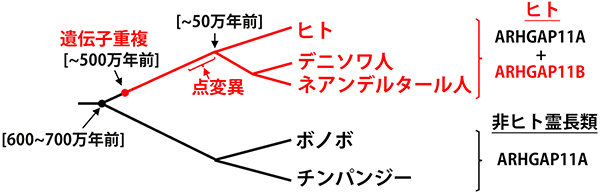
図2 系統図とARHGAP11Bの発生チンパンジーとヒトとの系譜が分岐したあとに、ARHGAP11Aの遺伝子重複が起こり、ARHGAP11Bが生じた。(発表論文内の図を改変)
研究の成果
本研究では、まず、ヒト特異的な遺伝子であるARHGAP11Bを、本来健常人で発現している生理的に近い量で発現するARHGAP11B遺伝子導入マーモセットを作製しました。脳の発生過程で、bRG細胞が豊富に存在して飛躍的に大脳新皮質が拡大する時期に注目し、胎生101日齢のコモンマーモセットの脳を解析しました。脳の解剖学的解析の結果、本来マーモセットではシワが存在しないはずの場所にも脳の凹凸が出現しており、野生型と比較してGI指数(脳の凹凸を示す指数)が約1.1倍に増高していました(図3)。次に、脳を非常に薄くスライスし、さまざまなタンパク質を各々特異的なマーカーでラベルして定量的な解析を行ったところ、 ARHGAP11B遺伝子導入マーモセットでは野生型に比べてbRG細胞の数が約4倍増加し、bRG細胞が多く存在する脳室下帯外側(図1-2)の厚さも約3.5倍厚くなっていることがわかりました。さらにbRGの増加に伴って、どこの神経細胞が増加しているのかを調べました。霊長類の進化に伴って増加する神経細胞は脳表に近い場所(皮質板浅層:図1-2)に多く存在しますが、ARHGAP11B遺伝子導入マーモセットではまさにそこにある神経細胞の数が約20%増加することがわかりました。
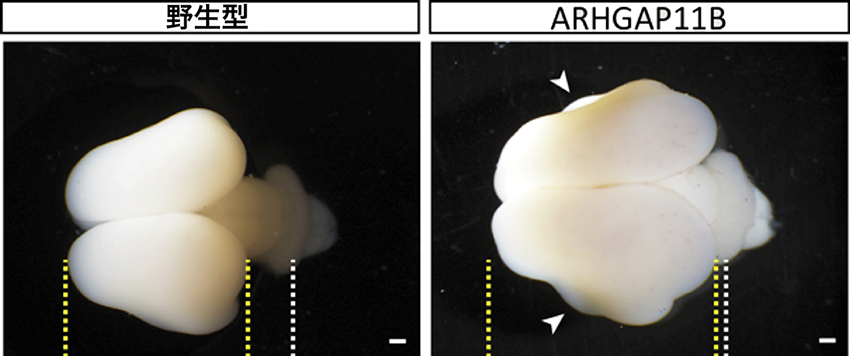 図3 ARHGAP11B導入による脳の拡大と凹凸の形成野生型およびARHGAP11B導入マーモセットの胎仔脳を頭頂方向から見たところ(胎生101日齢)。黄破線;大脳皮質の境界線。白破線;小脳の位置。矢印;本来ないはずの脳の凹凸。スケールバー=1mm。(発表論文内の図を改変)
図3 ARHGAP11B導入による脳の拡大と凹凸の形成野生型およびARHGAP11B導入マーモセットの胎仔脳を頭頂方向から見たところ(胎生101日齢)。黄破線;大脳皮質の境界線。白破線;小脳の位置。矢印;本来ないはずの脳の凹凸。スケールバー=1mm。(発表論文内の図を改変)
以上のことから、本研究を通じて、ヒトがチンパンジーから分岐したのちの大脳新皮質の拡大はARHGAP11Bによって引き起こされたことを、非ヒト霊長類モデルを用いて証明することに成功しました。
研究の意義と今後の展開
大脳新皮質(図1-1)は、大脳皮質構造のうち進化的に新しい部分であり、知覚、記憶、思考、随意運動、言語などをつかさどっています。ヒト特異的な遺伝子であるARHGAP11Bによってもたらされた新皮質の拡大は、ヒトに特徴的な新皮質の機能の獲得に関連している可能性が高いと考えられます。今後、ヒトだけが獲得し得た新皮質の機能の解明につながるかもしれません。
特記事項
本研究は、主にCentral funds from the Max Planck Societyおよび国立研究開発法人日本医療研究開発機構(AMED)革新的技術による脳機能ネットワークの全容解明プロジェクトの支援によって行われました。
論文
- 英文タイトル:
- Human-specific ARHGAP11B increases size and folding of primate neocortex in the fetal marmoset
- タイトル和訳:
- ヒトに特異的な遺伝子であるARHGAP11Bは非ヒト霊長類であるマーモセット胎仔脳の大脳新皮質を拡大させ、脳回脳溝構造を増加させる
- 著者名:
- Michael Heide、Christiane Haffner、村山綾子、黒滝陽子、篠原晴香、 岡野栄之、佐々木えりか、Wieland B. Huttner
- 掲載誌:
- Science
- DOI:
- 10.1126/science.abb4129
用語解説
- (注1)コモンマーモセット:
- 新世界ザルに分類され、ブラジル北東部原産の小型の非ヒト霊長類。体重は300~500g程度とラットぐらいの大きさで、耳の毛ぶさや長い尾を特徴とする。一夫一婦(もしくは二婦)制の家族単位で行動し、多種の鳴き声でコミュニケーションをとる。非ヒト霊長類のなかでも多産で、妊娠期間145日前後であり、繁殖に季節性はなく、1回に1~4頭を出産する。近年、非ヒト霊長類の実験動物として広く使用されており、遺伝子改変も可能である。
- (注2)脳回脳溝構造:
- 脳のシワの凸部分(脳回)と凹(溝)部分(脳溝)で構成される構造。
- (注3)bRG細胞:
- basal radial gliaの略。oRG(outer radial glia)細胞ともいう。神経幹細胞が生み出す幼弱な神経前駆細胞の中でも、神経幹細胞のように神経細胞を多く生み出す力を保持した、非常に未分化度の高い神経前駆細胞。胎生期の大脳皮質の外側脳室下帯という領域に存在し、多くの神経細胞を生み出す第2の細胞増殖帯を構成する。(図1-2)
- (注4)神経幹細胞:
- 自らと同じ性質を保ったまま分裂できる能力(自己複製能)と神経系の細胞種すべてに分化しうる能力(多分化能)を併せ持った、未だ細胞の運命決定が脆弱な状態(未分化度の高い)の神経系細胞。(図1-2)
- (注5)滑脳:
- 脳回脳溝構造(脳のシワ)がない脳。<—>(反意語)皺脳
- (注6)フェレット:
- イタチ科に属する哺乳類で、古くから家畜化されている。脳にシワがある。近年、実験動物としても盛んに使われている。
- (注7)部分的重複:
- 遺伝子を含むDNAのある領域が一部分だけ重複する現象。
お問い合わせ先
本発表資料のお問い合わせ先
慶應義塾大学医学部生理学教室
教授 岡野 栄之(おかの ひでゆき)
公益財団法人実験動物中央研究所 マーモセット医学生物学研究部
部長 佐々木 えりか(ささき えりか)
本リリースの発信元
慶應義塾大学
信濃町キャンパス総務課:鈴木・山崎
公益財団法人実験動物中央研究所
広報室
理化学研究所
広報室 報道担当
AMED事業に関すること
日本医療研究開発機構
疾患基礎研究事業部 疾患基礎研究課
革新的技術による脳機能ネットワークの全容解明プロジェクト

