2020-09-14 理化学研究所,日本医療研究開発機構
理化学研究所(理研)脳神経科学研究センター分子精神遺伝研究チームの島本(光山)知英研究員、吉川武男チームリーダーらの国際共同研究グループ※は、統合失調症[1]患者の脳の白質[2]において、脂質代謝に関連する遺伝子ネットワーク[3]の乱れを発見しました。
本研究成果は、一部の統合失調症患者で観察される白質病変の原因となるメカニズムの理解に役立つもので、新しい治療法開発の切り口になると期待できます。
今回、国際共同研究グループは、質量分析装置[4]を用いた統合失調症患者死後脳の脳梁[5](代表的な白質)の脂質定量解析により、一部の統合失調症患者の脳梁では、脂質組成(特定の脂質の含量パターン)が変動していることを発見しました。さらに、遺伝学的解析によりNFATC2[6]という転写因子[7]を起点とした遺伝子ネットワークと、グリア細胞の一つであるミクログリア[8]の異常が根底にあることを見いだしました。これらの結果から、NFATC2を起点とした遺伝子ネットワークとミクログリアの異常が関連した脂質組成変化が、統合失調症病態の形成に関与する可能性が示されました。
本研究は、科学雑誌『Cerebral CORTEX』の掲載に先立ち、オンライン版(2020年9月14日付:日本時間2020年9月14日)に掲載されます。

統合失調症患者の脳梁(白質)で変動していた遺伝子と各細胞マーカー遺伝子の相関関係
- ※国際共同研究グループ
- 理化学研究所 脳神経科学研究センター
分子精神遺伝研究チーム
チームリーダー 吉川武男(よしかわたけお)
副チームリーダー 大西哲生(おおにしてつお)
研究員 島本(光山)知英(しまもと・みつやまちえ)
研究員 シャビーシュ・バラン(Shabeesh Balan)
研究員 江崎加代子(えさきかよこ)
客員研究員 豊田倫子(とよたともこ)
キャリア形成推進プログラム
上級研究員 前川素子(まえかわもとこ)
研究基盤開発部門 生体物質分析ユニット
専門技術員 岩山佳美(いわやまよしみ)
東京大学大学院 新領域創成科学研究科 メディカル情報生命専攻
ゲノムデータサイエンス解析講座
特任教授 中谷明弘(なかやあきひろ)
The Florey Institute of Neuroscience and Mental Health, Howard Florey Laboratories, The University of Melbourne /The Centre for Mental Health, Swinburne University(オーストラリア)
教授 ブライアン・ディーン(Brian Dean) - 研究支援
- 本研究は、日本学術振興会(JSPS)科学研究費補助金若手研究(B)「精神疾患の病態生理に関与する機能性脂質の探索・同定(研究代表者:島本知英)」、同若手研究「統合失調症新規病態仮説「脂質生合成代謝異常-脳梁形成・機能不全仮説」の検証と応用(研究代表者:島本知英)」、同新学術領域研究「マルチスケール精神病態の構成的理解(領域代表者:林(高木)朗子)」、日本医療研究開発機構(AMED)革新的先端研究開発支援事業(AMED-CREST)『画期的医薬品等の創出をめざす脂質の生理活性と機能の解明』研究開発領域(研究開発総括:横山信治)における研究開発課題「光による脂質の同定制御観察技術すなわちオプトリピドミクスの創生(研究開発代表者:瀬藤光利)」、脳科学研究戦略推進プログラム『臨床と基礎研究の連携強化による精神・神経疾患の克服(融合脳)』の「細胞内代謝・ダイナミクス制御から切り拓く発達障害・統合失調症の病理の解明・新規治療法の開発(代表:吉川武男)」による支援を受けて行われました。
背景
幻覚や妄想、意欲の低下や認知機能障害などの症状を示す統合失調症は、発症頻度が一般人口の約1%であり、決して稀有な精神疾患ではありません。現在の治療法では、治療を行っても改善が見られない、いわゆる「治療抵抗性」の患者が全体の3割に上るという報告があり注1)、さらに薬の副作用に悩まされる患者も多くいます。こうした現状にもかかわらず、新しい治療薬の開発がなかなか進まない理由として、統合失調症の詳しい発症・病態メカニズムがよく分かっていないことが挙げられます。
これまでの統合失調症研究から、脂質(特に脂肪酸)が何らかの形で疾患の発症に影響を与えているという説が注目されてきました。近年、核磁気共鳴画像(MRI)[9]などの画像解析技術の進歩により、脳の中でも特に脂質に富む領域である白質において、容積の減少や神経線維の走行の異常、ミエリン[10]の構造異常が数多く報告されています。しかし、その原因となる分子メカニズムはよく分かっておらず、白質と脂質との関係性についてもあまり調べられていませんでした。
そこで国際共同研究グループは、白質の中でも左右の大脳半球をつなぐ大きな構造をしている脳梁における脂質代謝を、生化学・遺伝学の観点から包括的に調べることにしました。
注1)John M. Kane et.al. Clinical Guidance on the Identification and Management of Treatment-Resistant Schizophrenia. J Clin Psychiatry 80(2):18com12123, 2019
研究手法と成果
白質は、脳の中でも特に脂質に富む領域です。これまでに国際共同研究グループは、脳梁(代表的な白質領域)のスフィンゴ脂質[11]解析を行い、特定の脂質の変動について発表しました注2)。脂質には非常に多くの種類が存在し、過去の統合失調症の臨床研究では、赤血球膜や血漿などにおける脂肪酸[12]や脂肪酸を構造中に持つ脂質の含量が変化していることが、数多く報告されてきました。
そこで今回、統合失調症患者で観察される白質の異常と脂肪酸を構造中に持つ脂質分子の関係性を明らかにするため、質量分析装置を用いて、代表的な白質である脳梁における100種以上の脂質分子の含量を定量的に測定しました。その結果、一部の統合失調症患者では、脳梁の脂質組成(特定の脂肪酸側鎖を持つ脂質の含量パターン)が変化していることが分かりました(図1)。

図1 脂質含量パターンに基づくサンプルの分類ヒートマップの縦軸は、各サンプルにおける各脂質分子(右側)の含量パターン(マスの色は相対的な脂質含量を示す)を示しており、横軸は統合失調症サンプル(赤)または対照サンプル(緑)を示している。左側の統合失調症7サンプル(紫枠)は、その他のサンプルとは脂質の含量パターンが異なっていることから、病態に脂質が関与している統合失調症サブグループの一つである可能性がある。
次に、統合失調症の脳梁で脂質組成が変化する原因を調べるために、脂質代謝に関与するタンパク質をコードする遺伝子(脂質代謝関連遺伝子)の発現量を調べました。解析には、差を検出しやすくするため脂質解析に用いたサンプルを含む拡大サンプルを用いました。その結果、統合失調症群では、発症していない人(対照群)と比較し、10種の脂質代謝関連遺伝子の発現が有意に低下、もしくは低下傾向を示しました。さらに、これらの脂質代謝関連遺伝子の発現調節をしている可能性の高い転写因子について、その遺伝子発現量を調べたところ、7種の転写因子の遺伝子発現量が統合失調症群で変動していました。興味深いことに、これらの遺伝子の多くが、転写因子であるNFATC2(Nuclear Factor of Activated T Cells 2)を起点とする遺伝子ネットワークの下流にあり、相互に関連していることが分かりました(図2)。
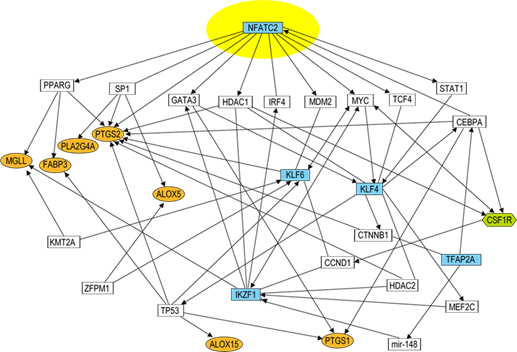
図2 NFATC2を起点とする遺伝子ネットワークこれまでの研究報告から判明している分子間の相互関係を示したネットワーク図。線はタンパク質間相互作用など、矢印は活性化やタンパク質-DNA相互作用、発現調節などを示す。統合失調症患者で変動が見られた転写因子(水色)および脂質関連遺伝子(オレンジ色)、ミクログリア関連遺伝子CSF1R(黄緑色)はNFATC2を起点とするネットワークの下流でつながっている。
脳は、神経細胞とグリア細胞(オリゴデンドロサイト[13]、アストロサイト[14]、ミクログリアなど)で構成されています。統合失調症群の脳梁で見られた脂質組成や遺伝子発現の変化と細胞種との関係を明らかにするため、各細胞のマーカーとなる遺伝子の発現を調べました。その結果、対照群と比較し、統合失調症群では、神経細胞、オリゴデンドロサイト、アストロサイトのマーカー遺伝子の発現量には差がありませんでしたが、ミクログリアのマーカー遺伝子の発現量が有意に低下していました(図3A)。そこで、ミクログリアの細胞密度調節に関与していることが知られている分子の一つであるCSF1R[15]をコードする遺伝子の発現について調べたところ、統合失調症群でCSF1R遺伝子の発現低下が見られました(図3B)。CSF1Rは、今回同定したNFATC2を起点とした遺伝子ネットワーク上の遺伝子と相互作用することが報告されています(図2)。
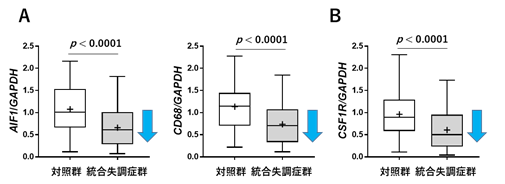
図3 統合失調症におけるミクログリア関連遺伝子の発現の異常A:統合失調症群では、ミクログリアのマーカー遺伝子AIF1(左図)、CD68(右図)の発現量が低下していた。
B:統合失調症群では、ミクログリアの密度制御に関与するCSF1Rをコードする遺伝子の発現量が低下していた。
次に、統合失調症群で変動していた10種の脂質代謝関連遺伝子および転写因子と各細胞マーカー遺伝子との発現量の相関関係を調べたところ、複数の遺伝子がミクログリアやオリゴデンドロサイトのマーカー遺伝子と正の相関を示しました(図4)。これらの結果から、ミクログリアの質的量的変化とオリゴデンドロサイトの質的変化が、統合失調症患者の脳梁で観察される脂質組成変動に関与している可能性が示されました。
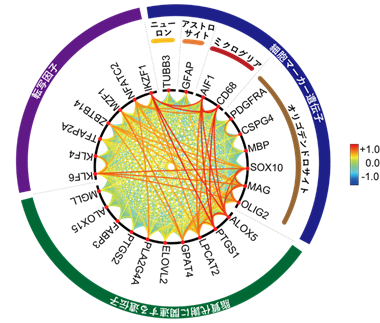
図4 統合失調症群の脳梁で変動していた遺伝子と各細胞マーカー遺伝子の相関関係図遺伝子同士をつなぐ線の色が赤く手前にあるほど、遺伝子間に強い正の相関関係があることを表している。複数の遺伝子が、ミクログリアのマーカー遺伝子やオリゴデンドロサイトのマーカー遺伝子と正に相関していることが分かる。
白質(脳梁)に豊富に存在するミエリンは、オリゴデンドロサイトによって形成されています。ミクログリアは、サイトカイン[16]などの情報伝達の役割を担う分子を介して、オリゴデンドロサイトと相互作用し、ミエリンの形成や維持などに関与しています。今回統合失調症患者の脳梁で発現低下していたNFATC2やCSF1Rが、ミクログリアのサイトカインの分泌に関与することから、国際共同研究グループは、統合失調症患者の脳梁ではミクログリアとオリゴデンドロサイト間の情報伝達に異常があるのではないかと考えました。そこで複数のサイトカインについて、それらをコードする遺伝子の発現量を調べたところ、2種類のサイトカインが統合失調症群で低下していることが分かり(図5)、さらにその発現量はNFATC2遺伝子やCSF1R遺伝子の発現と正に相関していました。これらの結果から、統合失調症患者の脳梁では、NFATC2を起点とした遺伝子ネットワーク異常が、ミクログリア-オリゴデンドロサイト間の情報伝達異常に関与している可能性が判明しました。またこの結果は、2019年に理研の吉川武男チームリーダーらが提案した、統合失調症では炎症反応(サイトカインの上昇)より抗炎症反応の方が本質的であることを支持しています注3)。
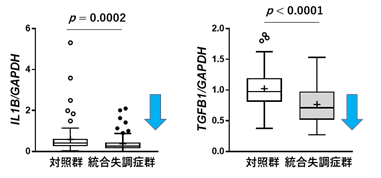
図5 統合失調症におけるサイトカインの遺伝子発現異常サイトカインであるIL1Bをコードする遺伝子(左図)とTGFB1をコードする遺伝子(右図)の発現量が統合失調症群で低下していた。
以上の結果から、転写因子NFATC2を起点した遺伝子ネットワークの異常とミクログリアの量的質的変化に関連した脂質組成異常が、統合失調症の病態と密接に関係している可能性が高いことが示唆されました(図6)。
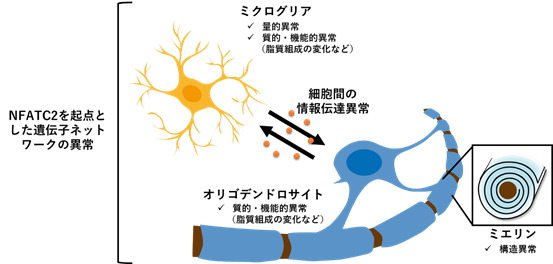
図6 NFATC2を起点とした遺伝子ネットワークおよびミクログリアの異常が引き起こす脂質組成変化と白質異常(仮説)
転写因子NFATC2を起点とした遺伝子ネットワークの異常が、ミクログリアの減少や機能異常(サイトカインの分泌能の低下など)を引き起こし、その結果、ミクログリア-オリゴデンドロサイト間の情報伝達、オリゴデンドロサイトの質的・機能的変化(脂質組成の変化など)が生じ、ミエリンの形成(構造異常)・維持に異常が生じるという仮説。
注2)2020年4月29日理化学研究所プレスリリース「統合失調症の脳で特定の脂質が低下」
注3)2019年10月28日理化学研究所プレスリリース「硫化水素の産生過剰が統合失調症に影響」
今後の期待
今回の研究により、一部の統合失調症の脳梁(白質)で脂質組成の変動が明らかになり、その原因としてNFATC2を起点とする遺伝子ネットワークやミクログリアの異常が関与している可能性が高いことが分かりました。この発見は、統合失調症の白質異常の根底にある遺伝子・分子・細胞の多層的なつながりを理解するための重要な手掛かりであるといえます。
NFATC2を起点とした遺伝子ネットワークとミクログリアの異常が関連する脂質組成変化が、白質の構造や機能、統合失調症の症状にどのような影響を与えるのかについては、今後Nfatc2遺伝子を破壊したマウスやミクログリアの細胞数を減少させたマウスなどを用いて、より詳細に解析する必要があります。
今回の研究結果は、一部の統合失調症患者で観察される白質病変の原因となるメカニズムの理解に役立つもので、転写因子をターゲットとした新たな治療抵抗性のある治療法開発の切り口になると期待できます。
論文情報
- タイトル
- Lipid Pathology of the Corpus Callosum in Schizophrenia and the Potential Role of Abnormal Gene Regulatory Networks with Reduced Microglial Marker Expression
- 著者名
- Chie Shimamoto-Mitsuyama, Akihiro Nakaya, Kayoko Esaki, Shabeesh Balan, Yoshimi Iwayama, Tetsuo Ohnishi, Motoko Maekawa, Tomoko Toyota, Brian Dean, Takeo Yoshikawa
- 雑誌
- Cerebral CORTEX
- DOI
- 10.1093/cercor/bhaa236
補足説明
- [1]統合失調症
- 代表的な精神疾患の一つであり、人口の約1%という比較的高い生涯発症率を持つ。幻覚や妄想などの「陽性症状」、意欲の低下や感情の平坦化などの「陰性症状」および「認知機能障害」が主要な症状で、社会的機能も低下する。現在の治療法では、治療効果が不十分であったり、再燃を繰り返すうちに次第に重篤化することも多い。そのため発症メカニズムの理解、およびそれに基づく根本的な治療法の開発が強く求められている。
- [2]白質
- 中枢神経組織の中で主に有髄神経線維が集積している部分。有髄神経線維のミエリンが生体膜の構成成分である脂質を豊富に含んでおり、白く見えることから、白質と呼ばれる。
- [3]遺伝子ネットワーク
- 遺伝子間の相互作用を表現するネットワークのこと。
- [4]質量分析装置
- 分子をイオン化し、その質量数と数を測定することにより、目的分子の同定や定量を行う装置。
- [5]脳梁
- 左右の大脳半球をつなぐ神経線維の巨大な束で構成される白質領域。
- [6]NFATC2
- 転写因子nuclear factor of activated T cells(NFAT)ファミリー分子の一つ。リン酸化により細胞質から核内に移行し、転写活性を示す。ミクログリアに高発現し、ミクログリアの機能調節に関与していることが報告されている。NFATC2はNuclear Factor Of Activated T Cells 2の略。
- [7]転写因子
- DNAに特異的に結合して、遺伝子の転写開始や転写調節に関与するタンパク質の総称。
- [8]ミクログリア
- 中枢神経系を構成するグリア細胞の一つ。脳内の免疫担当細胞として知られている。
- [9]核磁気共鳴画像(MRI)
- 磁力と電磁波の力を使って体内の状態を断面像として描写する検査。MRIはMagnetic Resonance Imagingの略。
- [10]ミエリン
- 神経軸索を渦巻き状に取り巻いている膜で、絶縁体として神経興奮伝達を助けている。中枢神経系では、オリゴデンドロサイトが多くの突起を出してミエリンを形成している。
- [11]スフィンゴ脂質
- スフィンゴ脂質は、スフィンゴイド塩基を骨格とした、スフィンゴ糖脂質やスフィンゴミエリンなどの脂質の総称。
- [12]脂肪酸
- 炭化水素の1価のカルボン酸のこと。炭素鎖に二重結合を持たない脂肪酸を飽和脂肪酸、二重結合を有する脂肪酸を不飽和脂肪酸という。脳では、アラキドン酸やドコサヘキサエン酸(DHA)などの二重結合を複数持つ多価不飽和脂肪酸が豊富に存在し、神経細胞の新生や分化、増殖など脳の発達に重要な役割を果たしている。また、脂肪酸は、細胞膜を構成するリン脂質やスフィンゴ脂質などの構成成分でもあり、その組み合わせにより脂質の構造多様性を生み出している。
- [13]オリゴデンドロサイト
- グリア細胞の一つ。オリゴデンドロサイトは一つの細胞から複数の樹状突起を伸ばし、複数の軸索に対してミエリン鞘を形成している。
- [14]アストロサイト
- グリア細胞の一つ。星型の形態をとることから星状膠細胞と呼ばれることもある。
- [15]CSF1R
- サイトカインであるコロニー刺激因子1(CSF1, colony stimulating factor 1)の受容体。ミクログリアに高発現し、ミクログリアの密度や機能調節に関与していることが知られている。CSF1Rはcolony stimulating factor 1 receptorの略。
- [16]サイトカイン
- 細胞から分泌されるタンパク質で、細胞間の情報伝達に関与する生理活性物質の総称。極めて微量で標的細胞を活性化したり、抑制したりする効果を持つことが知られている。
発表者・機関窓口
発表者
理化学研究所 脳神経科学研究センター 分子精神遺伝研究チーム
研究員 島本(光山)知英(しまもと(みつやま)ちえ)
チームリーダー 吉川武男(よしかわたけお)
機関窓口
理化学研究所 広報室 報道担当
AMED事業に関する問い合わせ先
日本医療研究開発機構(AMED)
シーズ開発・研究基盤事業部 革新的先端研究開発課

