2018/09/26 基礎生物学研究所
私たちが眼で物や景色を見ると、それらの像は眼の中の網膜と呼ばれる映画館のスクリーンのようなシート状の組織に映し出されます。これらの像の情報(視覚情報)は網膜でいったん処理された後(色や動きなどの要素に分解される)、脳内に伝えられます。視覚情報を脳に伝えているのが、網膜から脳内に長く伸びた視神経の軸索です。発生期においては、網膜から伸び出した視神経軸索は正しいルートを選択して脳内の目的領域に到達し、さらにその領域内で適切な相手を見つけ出して結合する(シナプス結合を作る)ことで神経回路を形成します。もし、正しい神経回路が作られないと、視覚に異常が生じます。
視神経の回路形成においては、Eph受容体と呼ばれる酵素(タンパク質のチロシンをリン酸化する酵素)が必須の役割を担っていることが明らかになっています。基礎生物学研究所 統合神経生物学研究部門の新谷隆史准教授と野田昌晴教授らは2006年に、ニワトリの視神経において、PTPROと呼ばれるタンパク質のチロシンに付いたリン酸をはずす酵素がEph受容体の働きを制御することによって、視神経の回路形成において重要な役割を果たしていることを明らかにしていました。
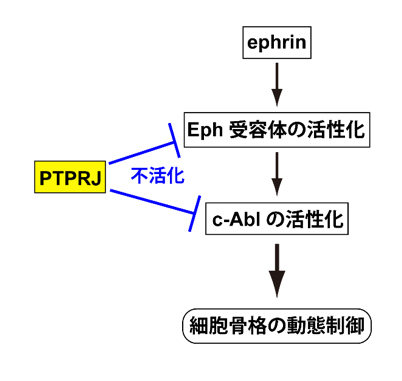
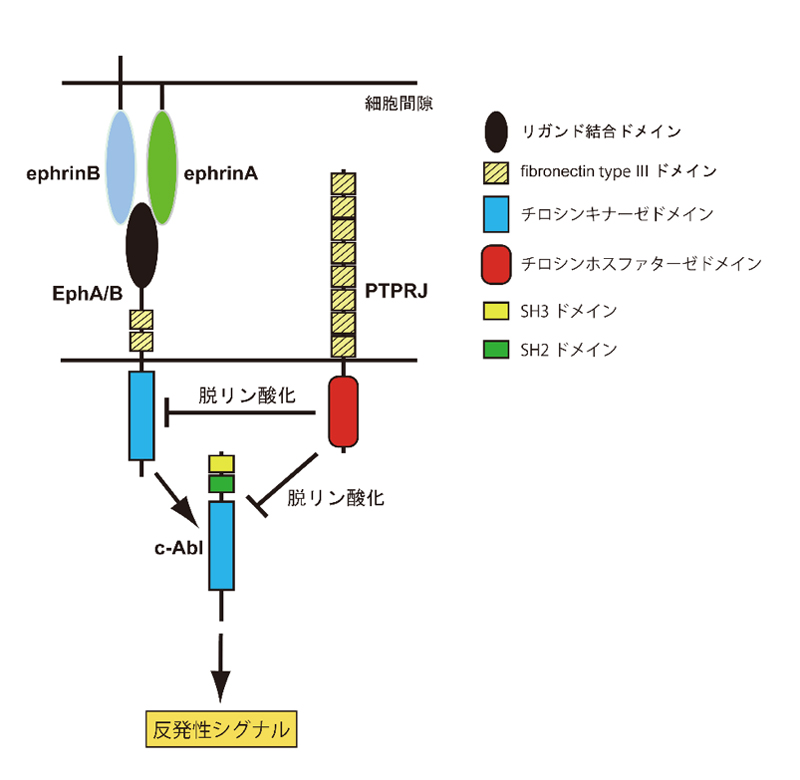
今回、同部門の于洋大学院生、新谷隆史准教授、及び野田昌晴教授らは、マウスでは、PTPROに代わってPTPRJがEph受容体の活性を制御していることを発見しました。研究グループはさらに、Eph受容体の反発性の情報伝達には、細胞内に分布しているc-Ablと呼ばれるタンパク質チロシンリン酸化酵素が必須の役割を果たしており、c-Ablの活性もPTPRJによって制御されていることを見出しました。すなわち、PTPRJはEph受容体とc-Ablの両者の活性を制御することにより、視神経の回路形成において重要な役割を果たしていました(下図参照)。
本成果はThe Journal of Neuroscience誌(2018年9月26日付)に掲載されます。
【研究の背景】
私たちの脳神経系が、感覚、運動、情動、記憶・学習などの高次の神経機能を発現するためには、発生過程において神経回路網が正しく形成されることが必要不可欠です。神経回路形成の研究において有用な系として用いられてきたのが、網膜から脳内の視中枢への視神経の投射系です。発生過程において、網膜から伸び出した視神経は視交叉を通過し、反対側の上丘においてトポグラフィックな投射(topographic projection)と呼ばれる非常に精緻な神経結合を形成します。トポグラフィックな投射とは、ある領域の神経細胞集団が二次元的な相対的位置関係を保った状態で他の神経細胞集団と神経結合を形成する、神経回路形成における基本様式の一つです。トポグラフィックな投射により、視覚情報は網膜に映し出された像の形状が保たれたままの状態で上丘に送られることになります。
視神経の投射の形成においては、Eph受容体と呼ばれる一群の受容体型タンパク質チロシンリン酸化酵素(RPTK)が必須の役割を担っていることが明らかになっています。Eph受容体は細胞外領域伝達ephrinと呼ばれるリガンド分子を認識して活性化し、主に反発性のシグナルを伝えます。すなわち、視交叉においてはephrinB2が高発現しており、このためEphB1を高発現する網膜の耳腹領域の視神経軸索は視交叉から反発作用を受けることにより、同側に投射します(図1A)。また、上丘においては、前側で低く後ろ側で高いという前後軸方向に沿ったephrinAの勾配が存在し、一方、網膜においては、鼻側で低く耳側で高いという鼻耳軸に沿ったEphAの勾配が存在しています。その結果、EphAを高発現する網膜耳側視神経は上丘の後ろ側に侵入できずに前側に投射し、EphAの発現が低い鼻側視神経は上丘の後ろ側に投射します(図1B)。
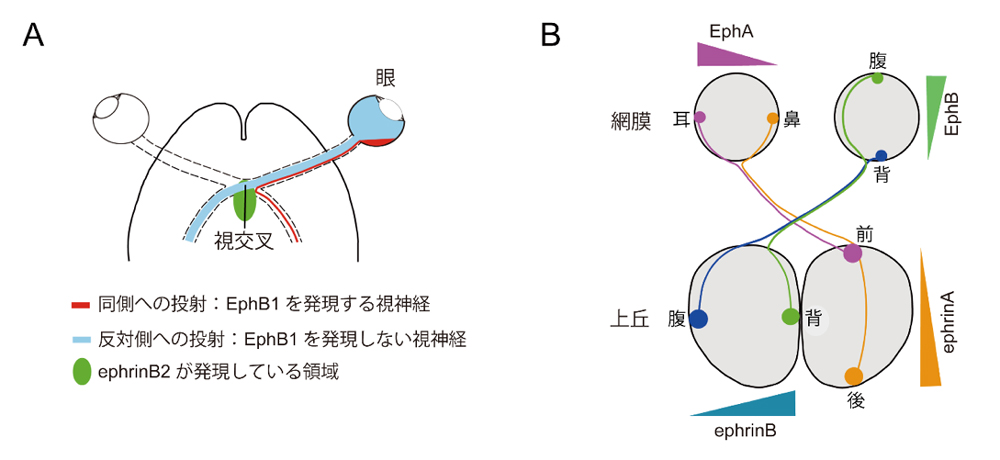
視交叉においてはephrinB2が高発現しており、このためEphB1を高発現する網膜の耳腹領域の視神経軸索は視交叉から反発作用を受けることにより、同側に投射する。残りのEphB1の発現が低い視神経は視交叉を通過して反対側に投射する。
(B)網膜から上丘へのトポグラフィックな投射。
網膜においては、鼻側で低く耳側で高いという鼻耳軸に沿ったEphAの勾配が存在している。
一方、上丘においては、前側で低く後ろ側で高いという前後軸方向に沿ったephrinAの勾配が存在する。EphAを高発現する網膜耳側視神経は上丘の後ろ側に侵入できずに前側に投射し、EphAの発現が低い鼻側視神経は上丘の後ろ側に投射する。背腹軸方向の投射にはEphBとephrinBによる誘因性の機構が働いている。
統合神経生物学研究部門では、これまでに様々な受容体様タンパク質チロシン脱リン酸化酵素(RPTP)の生理機能を明らかにする研究を進めてきました。RPTPはタンパク質のチロシンに付いたリン酸をはずす(脱リン酸化する)酵素で、細胞表面に存在しています。2007年に研究グループは、RPTPの一つであるPTPROがニワトリの視神経において、Eph受容体の活性を制御することによって、視神経のトポグラフィックな投射に重要な役割を果たしていることを見出しました(論文1)。一方、哺乳類であるマウスの視神経において、Eph受容体の活性を制御しているRPTPはこれまで不明でした。
【研究の成果】
研究グループは、2013年に培養細胞を用いた解析から、哺乳類のR3 PTPサブファミリーに属するPTPRO, PTPRJ, PTPRB, PTPRHのいずれもがEphを脱リン酸化することを明らかにしていました(論文2)。そこで、発生期のマウス脳膜におけるR3 PTPサブファミリーの発現を調べところ、PTPRJとPTPROの二つが網膜に発現しており、PTPRJの発現量がPTPROの発現量よりも顕著に高いことを見出しました。
次に、PTPRJとPTPROの視神経投射における役割を明らかにするために、それぞれの遺伝子欠損マウスと両者の二重欠損マウスについて解析を行いました。その結果、Ephのリン酸化は、野生型マウスに比べてPtprj欠損マウスと二重欠損マウスにおいて有意に高いことが分かりました(図2)。
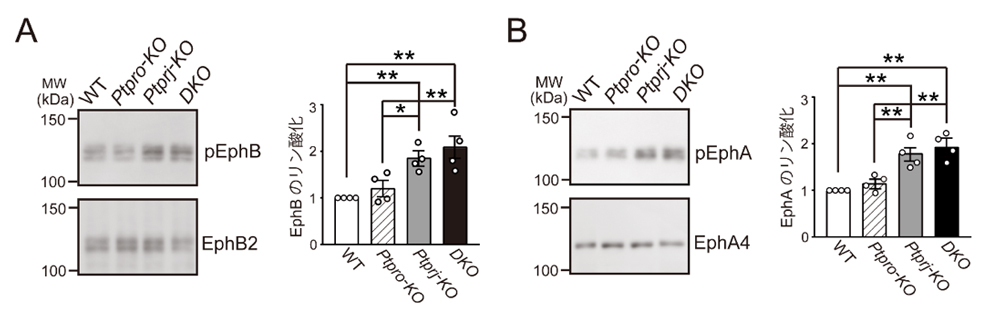
さらに、網膜の培養系を用いた解析によって、EphAとEphBを介した軸索の反発性の応答が野生型マウスに比べてPtprj欠損マウスと二重欠損マウスにおいて有意に増強していることが明らかになりました。以上の結果から、マウスの視神経では、PTPRJがEphの活性制御を行っていることが明らかになりました(図3)。

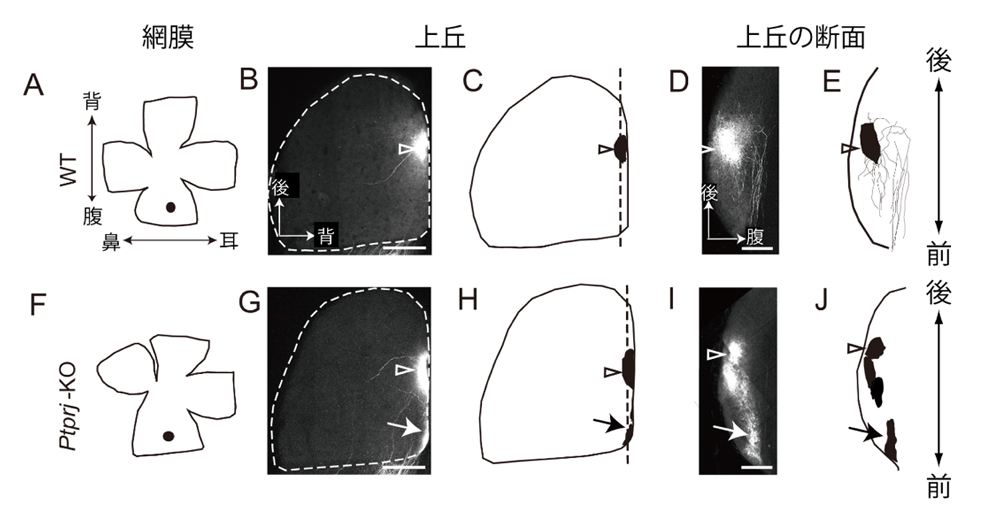
続いて、各マウスの視神経の投射について調べたところ、視交叉を避けて同側性に投射する視神経の割合と、反対側の眼に向かって逆走行する視神経の割合が野生型マウスに比べてPtprj欠損マウスと二重欠損マウスにおいて有意に増加していることが分かりました。また、上丘におけるトポグラフィックな投射について調べたところ、Ptprj欠損マウスと二重欠損マウスにおける視神経の投射位置が正常な投射位置から前側にシフトしていることが明らかになりました(図4)。これらの結果は、PTTPRJによるEphの活性抑制が無くなったために、視神経軸索がephrinに対してより強い反発性の応答をしたためと解釈されます。
研究グループはさらに、Eph受容体の反発性の情報伝達において、非受容体型のタンパク質チロシンリン酸化酵素であるc-Ablが必須の役割を果たしていることを明らかにしました。PTPRJはc-Ablを基質として脱リン酸化することによって、その活性化を抑制していました。これらの結果から、Ptprj欠損マウスにおいては、c-Ablが活性化しやすくなっていると考えられました。そこで、c-Ablを活性化する薬剤を発生期のマウスに投与したところ、Ptprj欠損マウスと二重欠損マウスにおいて観察されたと同様に、視交叉において同側性もしくは反対の眼に向かって投射する視神経の割合が増加するとともに、上丘における視神経の投射位置が前側にシフトすることが明らかになりました。以上の結果から、PTPRJはEph受容体とc-Ablの両方を基質としており、両者を脱リン酸化することによって反発性シグナルの制御に働いていることが明らかになりました。
【今後の展望】
本研究では、PTPRJがEphとc-Ablを基質として脱リン酸化することで、視神経の回路形成において重要な役割を果たしていることを明らかになりました。PTPRJとEph、c-Ablは視神経だけでなく、発生期及び成体の神経系において広く発現しています。このため、PTPRJはEphとc-Ablの活性を制御することにより、神経系の様々な領域の神経回路形成に働いていると予想されます。また、Ephとc-Ablは神経軸索の投射だけでなく、シナプスの形成や維持においても重要であることが分かっているため、PTPRJはこれらにも関与することが考えられます。今後は、このようなシナプスにおけるPTPRJの機能について解析することで、シナプスの形成と維持に関する新しい仕組みを明らかにできると期待しています。
【参考論文】
論文1:Shintani, T., Ihara, M., Sakuta, H., Takahashi, H., Watakabe, I., and Noda, M. (2006) Eph receptors are negatively controlled by protein tyrosine phosphatase receptor type O. Nature Neurosci. 9, 761-769.
基生研ニュース:リン酸化酵素の機能を制御するメカニズムを解明(2006.05.08)
http://www.nibb.ac.jp/pressroom/news/2006/05/post-121.html
論文2:Sakuraba, J., Shintani, T., Tani, S., and Noda, M. (2013) Substrate specificity of R3 receptor-like protein tyrosine phosphatase subfamily towards receptor protein tyrosine kinases. J. Biol. Chem. 288, 23421-23431.
基生研ニュース:R3 RPTPサブファミリーが多数のRPTKを基質にしていることを発見(2013.07.08) http://www.nibb.ac.jp/press/2013/07/08.html
【発表雑誌】
The Journal of Neuroscience 2018年9月26日号
論文タイトル:Protein Tyrosine Phosphatase Receptor Type J (PTPRJ) Regulates Retinal Axonal Projections by Inhibiting Eph and Abl Kinases in Mice
著者: Yang Yu, Takafumi Shintani, Yasushi Takeuchi, Takuji Shirasawa, and Masaharu Noda
【研究グループ】
本研究は、基礎生物学研究所・統合神経生物学研究部門の野田昌晴教授と新谷隆史准教授が中心となり実施されました。Ptpro遺伝子欠損マウスは東京都老人総合研究所の白澤卓二の研究室で作製され理化学研究所で維持されていたものを使用しました。
【研究サポート】
本研究は、文部科学省科学研究費助成事業の支援を受けて行われました。
【本研究に関するお問い合わせ先】
基礎生物学研究所 統合神経生物学研究部門
教授:野田 昌晴 (ノダ マサハル)
【報道担当】
基礎生物学研究所 広報室

