2021-04-08 東京大学
寺島 一郎(生物科学専攻 教授)
松尾 光敏(研究当時:生物科学専攻 修士課程2年生)
鈴木 祥弘(神奈川大学理学部生物科学科 准教授)
矢守 航(大学院農学生命科学研究科 附属生態調和農学機構 准教授)
河野 優(生物科学専攻 特任助教)
発表のポイント
- 薄暗い環境に耐える耐陰性植物のクワズイモの葉緑体は、ときおり当たる強光にも傷つきにくく、しかも、弱光で栽培した植物の葉緑体ほど耐性が高いことがわかりました。
- 定常的な強光には、強光で栽培した植物の葉緑体ほど耐性がありますが、木漏れ日が射し込む林の中のように光の強さが変動する場合には、逆に、弱い光で栽培する方が耐性が高まるのです。
- 強光による光合成器官の傷害(光阻害)は、植物の生産力を低下させる大きな要因の一つです。本研究の知見は、自然光条件下における強光傷害耐性植物の創出のための基盤となるでしょう。
発表概要
光合成は光のエネルギーを使って有機物を合成する反応で、葉の細胞に含まれている葉緑体で行われます。光は光合成に必須ですが、強すぎる光は葉緑体を損傷してしまいます。光のエネルギーを化学エネルギーに変換する光化学系 I と II(注1) のうち、強い光を当て続けたときにおもに損傷をうけるのは光化学系 II であること、強光で栽培した植物ほどこの損傷に耐性であることは、よく知られていました。
東京大学大学院理学系研究科の寺島一郎教授、松尾光敏大学院生(研究当時)、河野優特任助教、東京大学大学院農学研究科の矢守航准教授、神奈川大学理学部生物科学科の鈴木祥弘准教授らは、数種類の植物に変動光を与えて、損傷を比較したところ、弱光環境下でも生育する耐陰性(注2)の強いクワズイモの光化学系 I は損傷を受けにくく、しかも、栽培時の光強度が弱いほど光化学系I の「変動光」耐性が強いことを発見しました。(図1)

図1:木漏れ日が当たっているクワズイモ。東京大学構内の圃場において撮影
この原因を探索すべく、葉のクロロフィル蛍光、光化学系 I 反応中心の酸化還元状態、単離した葉緑体の種々のスペクトル、クロロフィルタンパク質複合体の組成などを解析しました。これらの結果、弱光栽培のクワズイモでは光化学系 I がよく励起されることや、強光下では光化学系 II 側から光化学系 I の反応中心に流れこむ電子の量が抑制されることが原因であることがわかりました。電子の流入量が少ないのによく励起されるので、光化学系 I の反応中心は電子を失った酸化状態となっていました。酸化された反応中心と還元された電子伝達体との電荷再結合(注3)により、過剰なエネルギーを消去していると考えられます。
光阻害は植物の光合成生産のネックとなっており、光阻害耐性植物の創出が試みられています。本研究で得られた知見は、自然条件下における光阻害耐性植物創出の鍵となるものです。
発表内容
光合成は光のエネルギーによって有機物を合成する反応です。植物や藻類の光合成産物は、 植物や藻類だけではなく、地球上のほとんどの生物を養っています。植物の光合成は、葉の細胞に含まれている葉緑体において行われます。光が葉緑体のタンパク質複合体に含まれるクロロフィルを励起すると、その励起状態はいくつかのクロロフィルを転移して、最終的に光化学系の反応中心クロロフィルに伝わります。反応中心クロロフィルが励起されると、電子が反応中心クロロフィルを飛び出し一連の酸化還元反応が開始されます。このように、光は光合成反応の駆動に必要不可欠なエネルギー源です。その一方で強すぎる光のエネルギーは、一重項酸素やスーパーオキシドなどの活性酸素(注4)の生成につかわれ、これらが葉緑体を損傷します。光化学系には、強い酸化力で水を酸化分解する光化学系 II と安定な還元剤をつくる光化学系 I とがあり、これまでに、定常的な強光による損傷を受けるのは光化学系 II であることがわかっていました。また、強光で栽培した植物ほど強光による損傷への耐性が強いことも明らかになっていました。
東京大学理学系研究科の河野優特任助教らは、野外の自然光環境でよく見られる、弱光に加 えて時々強光が当たるような「変動光」下では、光化学系 II ではなく I が損傷されることを見出し、この原因を追求してきました。光化学系 I のみを駆動する遠赤色光が存在すると、この阻害がほとんど起こらなくなることも見出しました。いくつかの種において、変動光による光化学系 I の損傷度合を比較したところ、弱光環境下でも生育できる耐陰性の強いクワズイモの光化学系 I は、遠赤色光がない場合にも変動光に耐性でした。しかも、弱光で栽培したクワズイモほど耐性が強いことがわかりました。(図2)
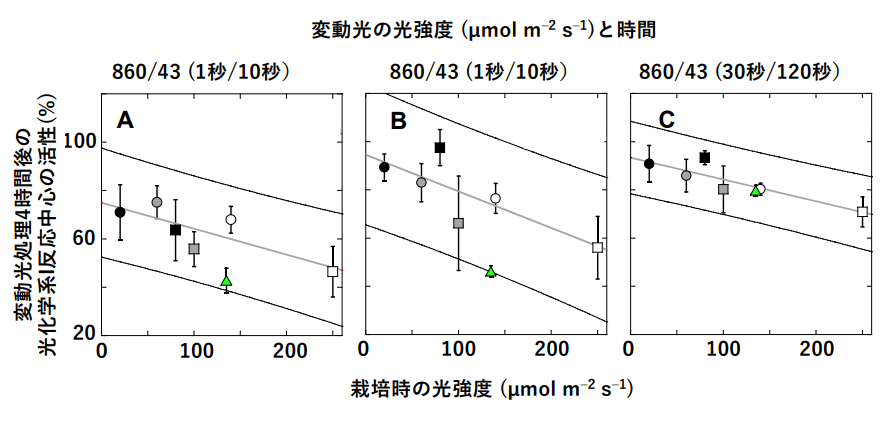
図2:変動光処理と栽培時の光強度が光化学系 I の活性に及ぼす影響。
強光と弱光とが交互に繰り返す 3 種類の変動光処理を 4 時間行った。100%光は栽培光よもかなり強く 860 µmol m-2 s-1の光合成有効光量子束密度(波長 400~700 nm の光量子が降ってくる量)である。これは真夏の昼間の光合成有効光量子束密度の 4 割程度である。植物の栽培は 12 時間の連続光で行った。光化学系 I の反応中心(P700)は光をあてると酸化される。図の縦軸には、変動光処理後の光によって酸化される P700 量の変動光処理前との比が示してある。各パネルの□と○で示したデータは、各光強度で栽培した 5 個体の葉の平均値と標準偏差。緑色の△は、モデル植物シロイヌナズナのデータである。クワズイモのデータに関する回帰直線と 95%信頼区間も示してある。
この原因を究明すべく、葉のクロロフィル蛍光や、光化学系 I 反応中心の酸化還元状態を精 査しました。また、単離した葉緑体についても種々のスペクトルを測定し、光化学系 I の反応 中心や電子伝達成分であるシトクロム f(注5)を定量しました。さらに、クロロフィルタンパク質複合体の組成も解析しました。これらから、弱光で栽培したクワズイモの光化学系 I が変動光による光阻害に耐性となる原因は以下の 2 つによることが明らかになりました。すなわち、1)弱光下で栽培したクワズイモでは、光化学系 II やシトクロム f の量に比べて光化学系 I を多く持っており、変動光の中の強光時に光化学系 II 側からシトクロム f を経て光化学系 I に流れこむ電子の量が少ないこと。2)強光時には、葉に吸収された光エネルギーが光化学系 I に多く分配されることです。光化学系 I への電子の流入量が少ないのに、光化学系 I に励起エネルギーが多く分配されると、光化学系 I の反応中心は電子を失った酸化状態となります。これと還元状態にある電子伝達コンポーネントとの電荷再結合によって、過剰なエネルギーがうまく消去されるようです。(図3)
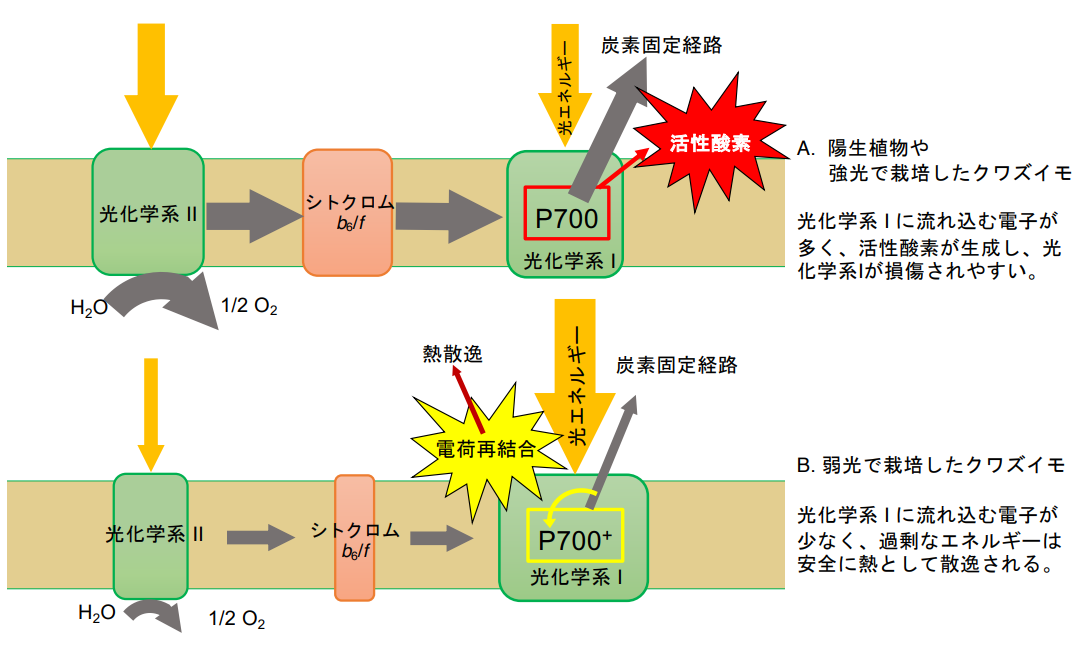
図3:変動光条件において弱光から強光になった時の光合成膜の様子。
A.陽生植物や強光下で栽培したクワズイモの場合:光化学系 II やシトクロム f を含むシトクロム複合体から光化学系 I に大量の電子が流れ込む。電子の流入量に対して光化学系 I への励起エネルギー量が十分でないので、光化学系 I の反応中心は還元状態(P700)となる。CO2固定経路駆動に必要な電子の量が光化学系 I に流れ込む電子の量よりも小さいと、電子は O2に流れてスーパーオキシドが形成し、これが原因となって光化学系 I が損傷される。
B.弱光下で栽培したクワズイモの場合:光化学系 II からシトクロム f を経て光化学系 Iに流れ込む電子フラックスが少なく、光化学系 I がよく励起される。光化学系 I の反応中心は酸化状態(P700+)となり、還元状態にある電子伝達成分と電荷再結合が起こる。電荷再結合の際、光化学系 I では過剰エネルギーは熱として安全に散逸されるため、光損傷が起こりにくい。
光阻害は植物の光合成生産を抑制しており、光阻害耐性植物の創出は作物の増産につながると考えられています。これまでに行われてきた光化学系 II に注目した研究だけではなく、光化学系 I の光阻害とその耐性の研究が必要であることを強く示すことができました。
発表雑誌
- 雑誌名
Photosynthesis Research論文タイトル
Photosystem I in low light-grown leaves of Alocasia odora, a shade-tolerant plant, is resistant to fluctuating light-induced photoinhibition著者
Ichiro Terashima*, Mitsutoshi Matsuo, Yoshihiro Suzuki, Wataru Yamori, Masaru Kono*DOI番号
https://doi.org/10.1007/s11120-021-00832-4
用語解説
注1 光化学系 I と II
光のエネルギーによってクロロフィルが励起され、その励起状態は、隣接するクロロフィル間を次々と転移し、最終的には反応中心とよばれるクロロフィルの二量体に到達する。反応中心クロロフィルが励起されると電子は反応中心を飛び出し、一連の電子伝達成分を還元する。電子を失った反応中心は強い酸化力を示す。光化学系 II はその酸化力で水を酸化し電子を奪う。光化学系 I からの電子は、水から得た電子によって NADP+を2 電子還元し、生化学反応に使われる安定な還元剤(NADPH)を生成する(NADP++H+ +2 e– →NADPH)。
注2 耐陰性
明るい場所を好む植物を陽生植物、薄暗い場所を好む植物を陰生植物とよぶ。クワズイモは、林縁などの明るい場所で花を咲かせるので陰生植物と呼んでしまうのには抵抗があるが、かなり暗い場所にも耐える耐陰性の強い植物で、インドア観葉植物として用いられる。モンステラ、ポトスなどと同じサトイモ科。
注3 電荷再結合
還元状態の光化学系 I の反応中心は 700 nm に吸収を示すので P700 と呼ばれる。酸化型のP700+と P700 近傍の還元状態にある電子伝達成分(A–)との間で電荷再結合がおこると、P700+ +A– → P700 + A となる。この時に生じるエネルギーが熱として散逸すれば、安全なアイドリングである。
注4 活性酸素
酸素(O2)が1電子還元されたスーパーオキシド(O2–)、励起されたクロロフィルが三重項状態となり、それが O2を励起した一重項酸素(シングレットオキシジェン、1O2)。両者とも酸化力が強く、生体物質を破壊する。
注5 シトクロム f
光化学系 II と I との間にある電子伝達物質。定常状態の光合成速度はいくつかの光合成コンポーネントによって同程度に律速される。その一つがシトクロムfで、定常強光で得られる最大光合成速度(一般には葉面積あたりで表す)は、シトクロム f の量と強い相関関係を示す。

