
マウス及びヒトなどの哺乳類において、脳由来神経栄養因子(BDNF)は、成熟個体の脳内で最も豊富に存在する神経栄養因子です。BDNFは、学習や記憶形成に深く関わっており、BDNFの分泌異常や受容体の機能障害は、心的外傷後ストレス障害(PTSD)、うつ病、認知機能障害等を引き起こすことが知られています。しかし、BDNFの産生制御に関わる分子機構は十分明らかにされていませんでした。基礎生物学研究所・統合神経生物学研究部門の鈴木亮子研究員と野田昌晴教授らの研究グループは、マウスの海馬で発現するSPIG1(SPARC-related protein containing immuno globulin domains 1)注1)が、BDNFの産生を抑制的に制御することによって、記憶の消去に関わっていることを明らかにしました。本研究成果は、Neurobiology of Learning and Memory(2018年7月号)に発表されます。(電子版は5月18日付で公開されました)。【研究の背景】
BDNFは、神経細胞内でBDNF前駆体として合成された後、分泌前あるいは分泌後にプロテアーゼにより切断を受けて、成熟体のBDNFになります(この過程をプロセシングと呼びます)。これまでに、このBDNFのプロセシングに関わるプロテアーゼはいくつか報告されましたが、プロセシングを調節する仕組みについては十分明らかにされていませんでした。2014年、同研究グループは、ニワトリ及びマウスを用いた研究から、 BDNFのプロセシングがSPIG1によって制御されていることを明らかにしました(Suzuki et al, J. Neurosci.34, 3429-3442, 2014)。また、Spig1遺伝子欠損マウス(生後10日目)の海馬において、神経スパイン注2)の数が野生型と比べて増加していること、さらに、成熟体のBDNF が多量に分泌され、それに応じて、受容体であるTrkB (Tropomyosin receptor kinase B )のリン酸化が亢進していることを明らかにしていました
(同上;2014年プレスリリース, http://www.nibb.ac.jp/press/2014/02/26.html)。【研究の成果】
記憶は、海馬の神経細胞間のシナプス(細胞間の接合部位)において、伝達効率が長期的に増加する現象(長期増強:LTP)が誘導される最初のステップを経て、形成されます。BDNFは、LTPを誘導する作用を有することが知られています。そこで、今回、小松由紀夫 名古屋大学名誉教授との共同研究により、Spig1遺伝子欠損マウスの海馬スライスにおけるシナプス伝達効率を調べました。その結果、Spig1遺伝子欠損マウスでは、野生型マウスに比べてLTPが亢進していることがわかりました(図1A)。さらに、BDNFの働きを抑える遮断抗体(anti-BDNF抗体)の添加によって、Spig1遺伝子欠損マウスで見られたLTPの亢進は、抑制されました(図1B)。次に、嫌悪記憶の形成後に、その記憶が消去される過程について調べました(図2)。Spig1遺伝子欠損マウスでは、野生型マウスに比べて記憶が速やかに消去されることが判明しました(図3A)。また、嫌悪記憶の消去学習を行った後、anti-BDNF抗体をマウスの脳室内に注入すると、その嫌悪記憶が一時的に回復することが判りました(図3B)。以上の結果は、SPIG1は、BDNFシグナルを通して記憶の消去に関わることを示しています。【成果の意義】
BDNFは、学習および記憶形成以外の生理機能(摂食抑制、糖代謝調節、心拍数や血圧の調節など)にも関わり、幅広い研究領域で研究されています。しかしながら、BDNFの生産調節のメカニズムはよく判っておりませんでした。本研究により、SPIG1がBDNFの成熟過程を制御することによって、記憶の消去に関与することが明らかになりました。過度な恐怖記憶の形成はPTSDを引き起こすことが知られており、本研究の知見は、PTSDの病態メカニズムの解明に役立つことが期待されます。
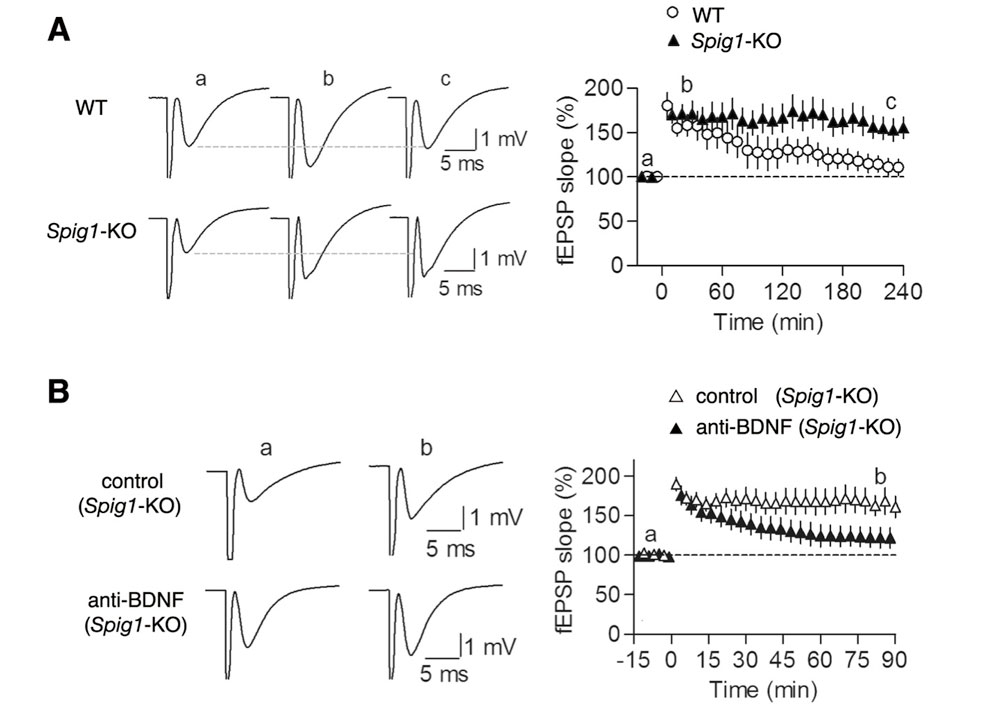 図1 Spig遺伝子欠損マウスにおけるLTP
図1 Spig遺伝子欠損マウスにおけるLTP
A, 野生型マウス(WT)およびSpig1遺伝子欠損マウス(Spig1-KO)の海馬CA1野スライスにおいて長期増強(LTP)を誘導する高頻度刺激を行い、興奮性シナプス後場電位(fEPSP, field excitatory postsynaptic potential)を測定した(a, -5分; b,+30分;c, +230分での測定結果; n = 6)。 Spig1遺伝子欠損マウスでは、LTPの亢進が観られた。
B, Spig1遺伝子欠損マウスの海馬CA1野スライスをanti-BDNF抗体で処理した後、高頻度刺激を行い、興奮性シナプス後場電位を測定した(a, -10分; b,+80分での測定結果; n = 6)。anti-BDNF抗体によって、LTPの亢進が抑制された。
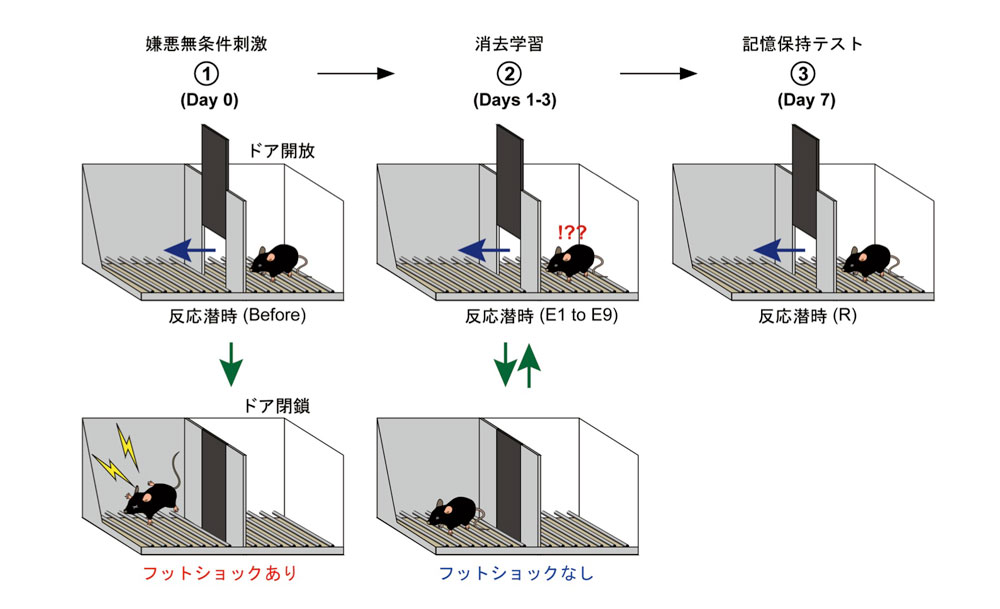 図2 嫌悪無条件刺激および嫌悪記憶の消去方法
図2 嫌悪無条件刺激および嫌悪記憶の消去方法
① 嫌悪無条件刺激 明暗の部屋をつくり、マウスを明室に置く。マウスは暗い環境を好むため短時間で、暗室へ移動する。マウスが暗室に入ったら、仕切り板を下げ、電気刺激(フットショック)を与える。
② 消去学習 フットショックを受けた翌日(Day1)に、明室に入れると、すくみ反応を示し、暗室に行くのを躊躇するが、日を経るに従って、早く暗室へ行くようになる(大丈夫であるということを学習する)。3日間、暗室へ入るまでの潜時(E1-E9)を測定する。
③ 記憶保持テスト 消去学習後7日目に、嫌悪記憶が消去されているかを調べる。暗室へ入るまでの潜時(R)を測定する。
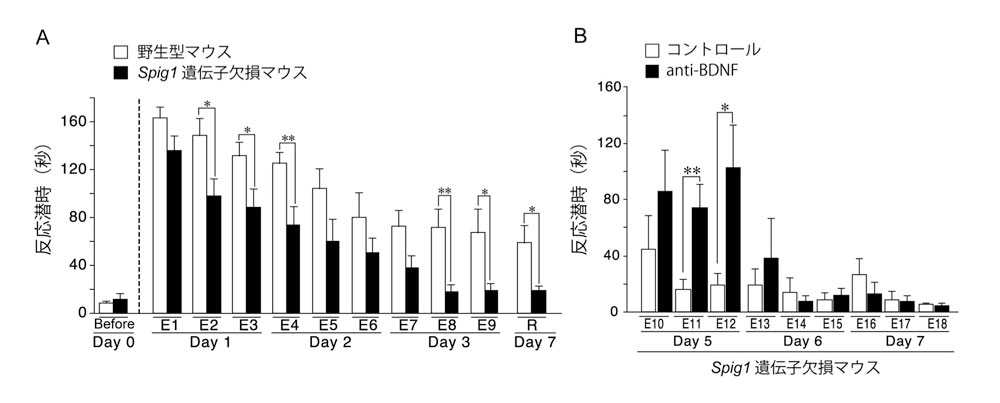 図3 嫌悪記憶の消去学習テスト
図3 嫌悪記憶の消去学習テスト
A, 嫌悪無条件刺激(フットショック)を受けたマウスに対して3日間(Day 1-3)、消去学習(E1-E9)を行い、明室から暗室へ移動するまでの潜時を測定した。Spig1遺伝子欠損マウスにおいては、嫌悪記憶が速やかに消去されていくのに対し、野生型マウスでは、嫌悪記憶が残った。7日後でも、両者の差は保持されていた (n = 12)。E, extinction training; R, memory retention。 *, P < 0.05, **, P < 0.01。
B, 消去学習を3日間行った翌日に、anti-BDNF抗体、あるいは、コントロール抗体を脳室内に注入した(Day 4)。その後、3日間(Day 5-7)、消去学習(E10-E18)を行った。anti-BDNF抗体を脳室内に注入したSpig1遺伝子欠損マウスでは、5日目に一時的に嫌悪記憶が回復した(n = 6)。 *, P < 0.05, **, P < 0.01。
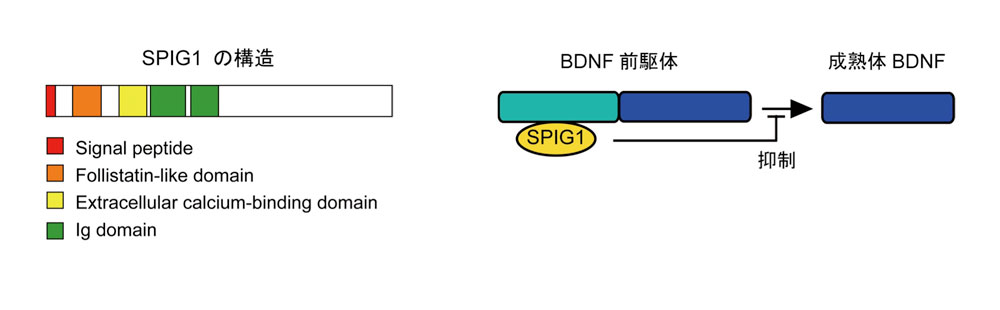
注1)SPIG1: SPIG1は、シグナルペプチド(赤)フォリスタチン様(プロテアーゼ阻害)ドメイン(橙)、細胞外カルシウム結合ドメイン(黄)、および免疫グロブリン様ドメイン(緑)から成る分泌因子である。BDNFの前駆体に結合し、BDNFのプロセシングを制御する。SPIG1の発現が抑制されると、成熟体のBDNFの量が増える。
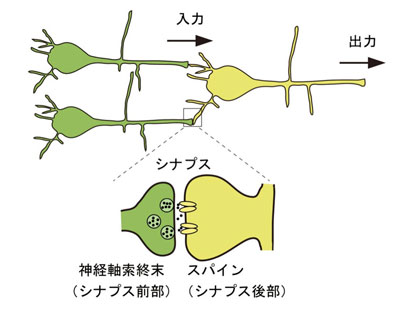
注2)スパイン:シナプス前部の神経細胞からシナプス後部の神経細胞が神経伝達物資を受け取る時のシナプス後部構造(棘突起)で、活動依存的に構造が変化するとされる。
Neurobiology of Learning and Memory 2018年5月18日電子版 (2018年7月号)
論文タイトル:Enhanced extinction of aversive memories in mice lacking SPARC-related protein containing immunoglobulin domains 1 (SPIG1/FSTL4)
著者:Ryoko Suzuki, Akihiro Fujikawa, Yukio Komatsu, Kazuya Kuboyama, Naomi Tanga and Masaharu Noda
https://doi.org/10.1016/j.nlm.2018.05.010【研究グループ】
本研究は、基礎生物学研究所・統合神経生物学研究部門の野田昌晴教授らの研究グループによって実施されました。【研究サポート】
本研究は、JSPS科学研究費助成事業 基盤研究(C)(18K06475)のサポートを受けて行われました。【本件に関するお問い合わせ先】
基礎生物学研究所 統合神経生物学研究部門
教授: 野田 昌晴 (ノダ マサハル)

